Back Clinic Клинична неврофизиологична поддръжка. Ел Пасо, Тексас. Chiropractor обсъжда д-р Александър Хименес клинична неврофизиология. Д-р Хименес ще изследва клиничното значение и функционалната активност на периферните нервни влакна, гръбначния мозък, мозъчния ствол и мозъка в контекста на висцерални и мускулно-скелетни нарушения. Пациентите ще придобият задълбочено разбиране за анатомията, генетиката, биохимията и физиологията на болката във връзка с различни клинични синдроми. Ще бъде включена хранителната биохимия, свързана с ноцицепцията и болката. И ще бъде подчертано прилагането на тази информация в терапевтичните програми.
Екипът ни се гордее с това, че предлага на нашите семейства и ранени пациенти само доказани протоколи за лечение. Като преподаваме цялостен холистичен уелнес като начин на живот, ние също променяме живота не само на нашите пациенти, но и на техните семейства. Правим това, за да можем да достигнем до колкото се може повече жители на Ел Пасо, които се нуждаят от нас, независимо от проблемите с достъпността. За отговори на всякакви въпроси, които може да имате, моля, обадете се на д-р Хименес на 915-850-0900.
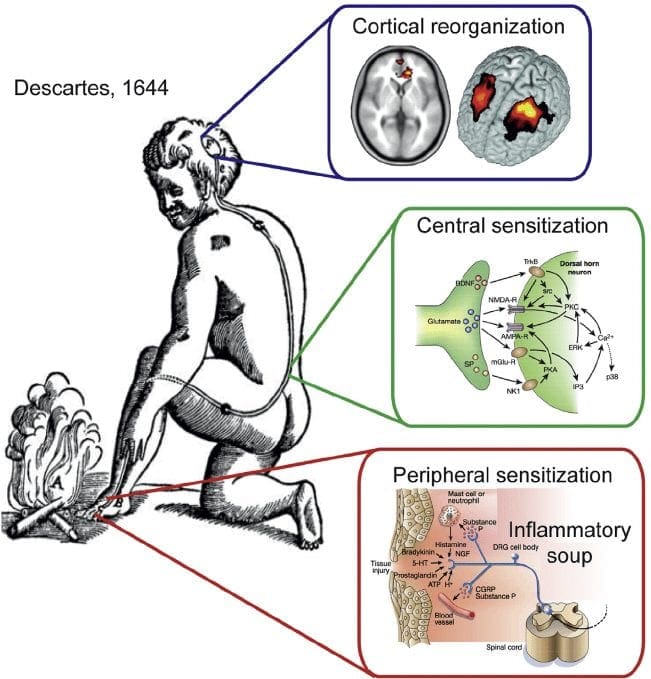
Невропатичната болка е сложно, хронично болково състояние, което обикновено е придружено от нараняване на меките тъкани. Невропатичната болка е често срещана в клиничната практика и също така представлява предизвикателство както за пациентите, така и за клиницистите. При невропатична болка самите нервни влакна могат да бъдат или повредени, дисфункционални или наранени. Невропатичната болка е резултат от увреждане от травма или заболяване на периферната или централната нервна система, където лезията може да се появи на всяко място. В резултат на това тези увредени нервни влакна могат да изпращат неправилни сигнали до други центрове за болка. Ефектът от нараняване на нервните влакна се състои в промяна в невронната функция, както в областта на нараняването, така и около нараняването. Клиничните признаци на невропатична болка обикновено включват сензорни феномени, като спонтанна болка, парестезии и хипералгезия.
Невропатичната болка, както е дефинирана от Международната асоциация за изследване на болката или IASP, е болка, инициирана или причинена от първична лезия или дисфункция на нервната система. Може да се получи от увреждане навсякъде по нервната ос: периферна нервна система, гръбначна или супраспинална нервна система. Чертите, които отличават невропатичната болка от други видове болка, включват болка и сетивни признаци, които продължават след периода на възстановяване. При хората се характеризира със спонтанна болка, алодиния или преживяване на безвредна стимулация като болезнена и каузалгия или постоянна пареща болка. Спонтанната болка включва усещания за „иглички“, парене, стрелба, пронизваща и пароксизмална болка или болка, подобна на електрически шок, често свързана с дизестезии и парестезии. Тези усещания променят не само сензорния апарат на пациента, но също така и благосъстоянието, настроението, вниманието и мисленето на пациента. Невропатичната болка се състои както от „отрицателни” симптоми, като загуба на сетивност и изтръпване, така и от „положителни” симптоми, като парестезии, спонтанна болка и повишено усещане за болка.
Състоянията, често свързани с невропатичната болка, могат да бъдат класифицирани в две големи групи: болка поради увреждане на централната нервна система и болка поради увреждане на периферната нервна система. Кортикални и субкортикални инсулти, травматични увреждания на гръбначния мозък, сирингомиелия и сирингобулбия, тригеминални и глософарингеални невралгии, неопластични и други заемащи пространство лезии са клинични състояния, които принадлежат към първата група. Невропатии при компресия или захващане на нерв, исхемична невропатия, периферни полиневропатии, плексопатии, компресия на нервните корени, след ампутационна пън и болка в фантомния крайник, постхерпетична невралгия и невропатии, свързани с рак, са клинични състояния, които принадлежат към последната група.
Патофизиология на невропатичната болка
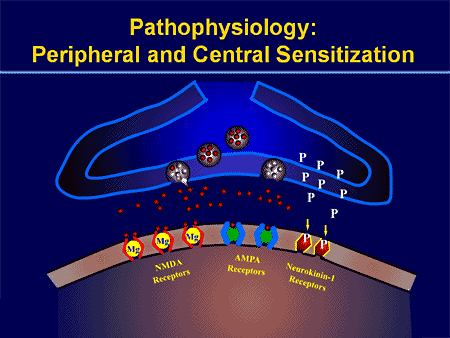
Патофизиологичните процеси и концепции, залегнали в основата на невропатичната болка, са множество. Преди да се обхванат тези процеси, прегледът на обикновените схеми за болка е от решаващо значение. Редовните вериги за болка включват активиране на ноцицептор, известен също като болков рецептор, в отговор на болезнена стимулация. Вълна на деполяризация се доставя до невроните от първи ред, заедно с натрий, който се втурва през натриеви канали и изтича калий. Невроните завършват в мозъчния ствол в тригеминалното ядро или в дорзалния рог на гръбначния мозък. Именно тук знакът отваря зависими от напрежението калциеви канали в предсинаптичния терминал, позволявайки на калция да влезе. Калцият позволява на глутамата, възбуждащ невротрансмитер, да бъде освободен в синаптичната област. Глутаматът се свързва с NMDA рецепторите на невроните от втори ред, причинявайки деполяризация.
Тези неврони преминават през гръбначния мозък и пътуват до таламуса, където синапсират с неврони от трети ред. След това те се свързват с лимбичната система и мозъчната кора. Съществува и инхибиторен път, който предотвратява предаването на сигнал за болка от дорзалния рог. Антиноцицептивните неврони произхождат от мозъчния ствол и се движат надолу по гръбначния мозък, където синапсират с къси интерневрони в дорзалния рог чрез освобождаване на допамин и норепинефрин. Интерневроните модулират синапса между неврона от първи ред, както и неврона от втори ред, като освобождават гама аминомаслена киселина или GABA, инхибиторен невротрансмитер. Следователно, спирането на болката е резултат от инхибиране на синапсите между неврони от първи и втори ред, докато усилването на болката може да бъде резултат от потискане на инхибиторните синаптични връзки.
Механизмът, който стои в основата на невропатичната болка, обаче, не е толкова ясен. Няколко проучвания върху животни разкриха, че могат да бъдат включени много механизми. Трябва обаче да се помни, че това, което се отнася за съществата, може да не винаги се отнася за хората. Невроните от първи ред могат да увеличат изстрелването си, ако са частично повредени и да увеличат количеството на натриеви канали. Ектопичните разряди са следствие от засилена деполяризация на определени места във влакното, което води до спонтанна болка и болка, свързана с движението. Инхибиторните вериги могат да бъдат намалени на нивото на дорзалния рог или мозъчните стволови клетки, както и на двете, позволявайки на импулсите на болката да пътуват без противопоставяне.
В допълнение, може да има промени в централната обработка на болката, когато поради хронична болка и употребата на някои лекарства и/или лекарства невроните от втори и трети ред могат да създадат „памет“ за болка и да станат сенсибилизирани. След това има повишена чувствителност на гръбначните неврони и намалени прагове на активиране. Друга теория демонстрира концепцията за симпатиковата невропатична болка. Тази идея е демонстрирана чрез аналгезия след симпатектомия от животни и хора. Въпреки това, комбинация от механика може да бъде включена в много хронични невропатични или смесени соматични и невропатични болкови състояния. Сред тези предизвикателства в областта на болката и много повече, що се отнася до невропатичната болка, е способността да се провери. Това има двоен компонент: първо, оценка на качеството, интензивността и напредъка; и второ, правилно диагностициране на невропатичната болка.
Има обаче някои диагностични инструменти, които могат да помогнат на клиницистите при оценката на невропатичната болка. Като начало, изследванията на нервната проводимост и сензорно предизвиканите потенциали могат да идентифицират и количествено определят степента на увреждане на сензорните, но не и ноцицептивните пътища чрез наблюдение на неврофизиологичните отговори на електрически стимули. Освен това, количественото сензорно тестване предприема стъпки за възприемане в реакция на външни стимули с различна интензивност чрез прилагане на стимулация върху кожата. Механичната чувствителност към тактилни стимули се измерва със специализирани инструменти, като например власинки на фон Фрей, убождане с преплетени игли, както и чувствителност към вибрации заедно с вибраметри и термична болка с термоди.
Също така е изключително важно да се извърши цялостна неврологична оценка за идентифициране на двигателни, сензорни и вегетативни дисфункции. В крайна сметка съществуват множество въпросници, използвани за разграничаване на невропатичната болка от ноцицептивната болка. Някои от тях включват само запитвания за интервю (напр. Невропатичният въпросник и ID Pain), докато други съдържат както въпроси за интервю, така и физически тестове (напр. скалата за оценка на невропатичните симптоми и признаци на Лийдс) и точния нов инструмент, Стандартизираната оценка на Болка, която съчетава шест въпроса за интервю и десет физиологични оценки.
Методи за лечение на невропатична болка
Фармакологичните схеми са насочени към механизмите на невропатичната болка. Въпреки това, както фармакологичните, така и нефармакологичните лечения осигуряват пълно или частично облекчение при само около половината от пациентите. Много основани на доказателства препоръки предполагат използването на смеси от лекарства и/или лекарства, за да функционират възможно най-много механизми. Повечето проучвания са изследвали предимно постхерпетична невралгия и болезнени диабетни невропатии, но резултатите може да не се отнасят за всички състояния на невропатична болка.
Антидепресанти
Антидепресантите повишават нивата на синаптичния серотонин и норепинефрин, като по този начин засилват ефекта на низходящата аналгетична система, свързана с невропатичната болка. Те са били основата на невропатичната болкова терапия. Аналгетичните действия могат да се дължат на блокада на обратното захващане на нор-адреналин и допамин, които вероятно повишават низходящото инхибиране, антагонизма на NMDA-рецепторите и блокадата на натриевите канали. Трициклични антидепресанти, като TCAs; например амитриптилин, имипрамин, нортриптилин и доксепин, са мощни срещу продължителна болка или пареща болка, заедно със спонтанна болка.
Трицикличните антидепресанти са доказано значително по-ефективни при невропатична болка от специфичните инхибитори на обратното захващане на серотонин или SSRI, като флуоксетин, пароксетин, сертралин и циталопрам. Причината може да е, че те инхибират обратното захващане на серотонин и нор-епинефрин, докато SSRI инхибират само обратното захващане на серотонин. Трицикличните антидепресанти могат да имат неприятни странични ефекти, включително гадене, объркване, блокове на сърдечната проводимост, тахикардия и камерни аритмии. Те могат също да причинят наддаване на тегло, намален праг на гърчове и ортостатична хипотония. Трицикличните лекарства трябва да се използват с повишено внимание при възрастни хора, които са особено уязвими към техните остри странични ефекти. Концентрацията на лекарството в кръвта трябва да се следи, за да се избегне токсичност при пациенти, които метаболизират бавно лекарства.
Инхибиторите на обратното захващане на серотонин-норепинефрин или SNRIs са нов клас антидепресанти. Подобно на TCAs, те изглежда са по-ефективни от SSRI за лечение на невропатична болка, тъй като те също инхибират обратното захващане както на нор-епинефрин, така и на допамин. Венлафаксин е толкова ефективен срещу инвалидизиращи полиневропатии, като болезнена диабетна невропатия, колкото имипрамин, като се споменава TCA, и двете са значително по-големи от плацебо. Подобно на TCAs, SNRIs изглежда дават ползи независимо от техните антидепресантни ефекти. Страничните ефекти включват седация, объркване, хипертония и синдром на отнемане.
Антипилептични лекарства
Антиепилептичните лекарства могат да се използват като лечение от първа линия, особено за някои видове невропатична болка. Те действат чрез модулиране на волтаж-зависимите калциеви и натриеви канали, чрез подобряване на инхибиторните ефекти на GABA и чрез инхибиране на възбуждащата глутаминергична трансмисия. Не е доказано, че антиепилептичните лекарства са ефективни при остра болка. При случаи на хронична болка антиепилептичните лекарства изглеждат ефективни само при тригеминална невралгия. Карбамазепинът се използва рутинно за това състояние. Известно е също, че габапентинът, който действа чрез инхибиране на функцията на калциевите канали чрез агонистични действия в алфа-2 делта субединицата на калциевия канал, е ефективен при невропатична болка. Габапентин обаче действа централно и може да причини умора, объркване и сънливост.
Неопиоидни аналгетици
Липсват силни данни, подкрепящи използването на нестероидни противовъзпалителни лекарства или НСПВС за облекчаване на невропатичната болка. Това може да се дължи на липсата на възпалителен компонент за облекчаване на болката. Но те са били използвани взаимозаменяемо с опиоиди като помощни средства при лечение на болка при рак. Въпреки това, има съобщения за усложнения, особено при тежко изтощени пациенти.
Опиоидни аналгетици
Опиоидните аналгетици са обект на много дебати за облекчаване на невропатичната болка. Те действат чрез инхибиране на централните възходящи импулси на болка. Традиционно невропатичната болка е била наблюдавана по-рано като опиоидна резистентност, при което опиоидите са по-подходящи методи за коронарни и соматични ноцицептивни видове болка. Много лекари предотвратяват използването на опиоиди за лечение на невропатична болка, до голяма степен поради опасения относно злоупотребата с наркотици, пристрастяването и регулаторните проблеми. Но има много проучвания, които са установили, че опиоидните аналгетици са успешни. Оксикодонът е по-добър от плацебо за облекчаване на болката, алодиния, подобряване на съня и увреждания. Опиоидите с контролирано освобождаване, според графика, се препоръчват за пациенти с постоянна болка, за да насърчат постоянни нива на аналгезия, да предотвратят колебанията в кръвната захар и да предотвратят нежелани събития, свързани с по-висока доза. Най-често се използват перорални препарати поради тяхната по-голяма лекота на употреба и рентабилност. Трансдермални, парентерални и ректални препарати обикновено се използват при пациенти, които не понасят перорални лекарства.
Местни анестетици
Близко действащите анестетици са привлекателни, защото благодарение на регионалното си действие имат минимални странични ефекти. Те действат чрез стабилизиране на натриевите канали в аксоните на периферните неврони от първи ред. Те работят най-добре, ако има само частично увреждане на нерва и са събрани излишни натриеви канали. Локалният лидокаин е най-добре проученият представител на курса за невропатична болка. По-конкретно, използването на този 5% лидокаинов пластир за пост-херпетична невралгия предизвика одобрението му от FDA. Пластирът изглежда работи най-добре, когато има увредена, но поддържана ноцицепторна функция на периферната нервна система от засегнатия дерматом, демонстрираща се като алодиния. Трябва да се постави директно върху симптоматичната област за 12 часа и да се елиминира за още 12 часа и може да се използва в продължение на години по този начин. Освен локални кожни реакции, той често се понася добре от много пациенти с невропатична болка.
Различни лекарства
Доказано е, че клонидин, алфа-2-агонист, е ефективен при подгрупа пациенти с диабетна периферна невропатия. Установено е, че канабиноидите играят роля в експерименталната модулация на болката при животински модели и доказателствата за ефикасността им се натрупват. СВ2-селективните агонисти потискат хипералгезията и алодинията и нормализират ноцицептивните прагове, без да предизвикват аналгезия.
Интервенционално управление на болката
Може да се обмисли инвазивно лечение за пациенти, които имат непреодолима невропатична болка. Тези лечения включват епидурални или периневрални инжекции на локални анестетици или кортикостероиди, имплантиране на епидурални и интратекални методи за доставяне на лекарства и поставяне на стимулатори на гръбначния мозък. Тези подходи са запазени за пациенти с непреодолима хронична невропатична болка, които са претърпели неуспешно консервативно медицинско лечение и също са претърпели задълбочена психологическа оценка. В проучване на Kim et al е показано, че стимулаторът на гръбначния мозък е ефективен при лечение на невропатична болка от произход на нервните корени.
Прозрение на д-р Алекс Химензе
При невропатичната болка се появяват симптоми на хронична болка, дължащи се на увредени, дисфункционални или наранени нервни влакна, обикновено придружени от увреждане или нараняване на тъканите. В резултат на това тези нервни влакна могат да започнат да изпращат неправилни сигнали за болка към други части на тялото. Ефектите от невропатичната болка, причинена от наранявания на нервните влакна, включват модификации в нервната функция както на мястото на нараняване, така и в областите около нараняването. Разбирането на патофизиологията на невропатичната болка е цел за много здравни специалисти, за да определят ефективно най-добрия подход за лечение, който да помогне за управлението и подобряването на нейните симптоми. От употребата на лекарства и/или лекарства, до хиропрактика, упражнения, физическа активност и хранене, могат да се използват различни подходи за лечение, за да се помогне за облекчаване на невропатичната болка за нуждите на всеки индивид.
Допълнителни интервенции за невропатична болка
Много пациенти с невропатична болка търсят допълнителни и алтернативни възможности за лечение за лечение на невропатична болка. Други добре познати схеми, използвани за лечение на невропатична болка, включват акупунктура, перкутанна електрическа нервна стимулация, транскутанна електрическа нервна стимулация, когнитивно поведенческо лечение, степенувани двигателни образи и поддържащо лечение и упражнения. Сред тях обаче хиропрактиката е добре познат подход за алтернативно лечение, който обикновено се използва за подпомагане на лечението на невропатична болка. Хиропрактиката, заедно с физиотерапия, упражнения, хранене и промени в начина на живот могат в крайна сметка да предложат облекчение на симптомите на невропатична болка.
Chiropractic Красота
Това, което се знае, е, че цялостното приложение за управление е от решаващо значение за борба с ефектите от невропатичната болка. По този начин хиропрактиката е програма за цялостно лечение, която може да бъде ефективна за предотвратяване на здравословни проблеми, свързани с увреждане на нервите. Хиропрактиката предоставя помощ на пациенти с много различни състояния, включително тези с невропатична болка. Страдащите от невропатична болка често използват нестероидни противовъзпалителни лекарства или НСПВС, като ибупрофен, или тежки болкоуспокояващи, отпускани с рецепта, за да облекчат невропатичната болка. Те могат да осигурят временно решение, но се нуждаят от постоянна употреба за управление на болката. Това неизменно допринася за вредни странични ефекти и в екстремни ситуации, зависимост от лекарства, отпускани по лекарско предписание.
Хиропрактиката може да помогне за подобряване на симптомите на невропатична болка и да подобри стабилността без тези недостатъци. Подход като хиропрактика предлага индивидуализирана програма, предназначена да определи първопричината за проблема. Чрез използването на гръбначни корекции и ръчни манипулации, хиропрактикът може внимателно да коригира всякакви изкривявания на гръбначния стълб или сублуксации, открити по дължината на гръбначния стълб, което може да намали последствията от нервно счупване чрез преместване на гръбначния стълб. Възстановяването на целостта на гръбначния стълб е от съществено значение за поддържането на високо функционираща централна нервна система.
Хиропрактикът може да бъде и дългосрочно лечение за подобряване на цялостното ви благосъстояние. Освен корекции на гръбначния стълб и ръчни манипулации, хиропрактикът може да предложи хранителни съвети, като например да предпише диета, богата на антиоксиданти, или може да разработи програма за физическа терапия или упражнения за борба с обострянията на нервната болка. Дългосрочното състояние изисква дългосрочно лечение и в това си качество медицински специалист, който е специализиран в наранявания и/или състояния, засягащи мускулно-скелетната и нервната система, като лекар по хиропрактика или хиропрактик, може да бъде безценен, докато работи за да прецените благоприятната промяна във времето.
Доказано е, че техниките за физическа терапия, упражнения и представяне на движение са полезни за лечение на невропатична болка. Хиропрактиката предлага и други методи на лечение, които могат да бъдат полезни за управлението или подобряването на невропатичната болка. Лазерната терапия на ниско ниво или LLLT, например, придоби огромна популярност като лечение на невропатична болка. Според различни изследователски проучвания се стигна до заключението, че LLLT има положителни ефекти върху контрола на аналгезията при невропатична болка, но са необходими допълнителни изследователски проучвания, за да се дефинират протоколи за лечение, които обобщават ефектите от ниско ниво на лазерна терапия при лечение на невропатична болка.
Хиропрактиката включва също хранителни съвети, които могат да помогнат за контролиране на симптомите, свързани с диабетна невропатия. По време на изследователско проучване е доказано, че диета с ниско съдържание на мазнини на растителна основа подобрява гликемичния контрол при пациенти с диабет тип 2. След около 20 седмици от пилотното проучване, участниците съобщават за промени в телесното си тегло и се съобщава, че електрохимичната проводимост на кожата в стъпалото се е подобрила с интервенцията. Изследователското проучване предполага потенциална стойност в интервенцията на нискомаслена растителна диета за диабетна невропатия. Освен това, клиничните проучвания установиха, че пероралното приложение на магнезиев L-треонат е в състояние да предотврати, както и да възстанови дефицитите на паметта, свързани с невропатична болка.
Хиропрактиката може също да предложи допълнителни стратегии за лечение за насърчаване на регенерацията на нервите. Например, се предполага, че подобряването на регенерацията на аксоните помага за подобряване на функционалното възстановяване след увреждане на периферните нерви. Установено е, че електрическата стимулация, заедно с упражнения или физически дейности, насърчава регенерацията на нервите след забавено възстановяване на нервите при хора и плъхове, според последните проучвания. Както електрическата стимулация, така и упражненията в крайна сметка бяха определени като обещаващи експериментални лечения за увреждане на периферните нерви, които изглеждат готови да бъдат прехвърлени към клинична употреба. Може да са необходими допълнителни изследвания, за да се определят напълно ефектите от тях при пациенти с невропатична болка.
Заключение
Невропатичната болка е многостранно образувание без конкретни насоки, за които да се грижим. Най-добре се управлява с помощта на мултидисциплинарен подход. Управлението на болката изисква текуща оценка, обучение на пациентите, осигуряване на проследяване на пациента и успокоение. Невропатичната болка е хронично състояние, което прави избора за най-добро лечение предизвикателен. Индивидуализиращото лечение включва отчитане на въздействието на болката върху благосъстоянието на индивида, депресията и уврежданията, заедно с продължаващо обучение и оценка. Изследванията на невропатичната болка, както на молекулярно ниво, така и при животински модели, са сравнително нови, но много обещаващи. Очакват се много подобрения в основните и клинични области на невропатичната болка, като по този начин се отварят вратите за подобрени или нови методи на лечение на това инвалидизиращо състояние. Обхватът на нашата информация е ограничен до хиропрактика, както и до наранявания и състояния на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителни теми: Болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати дни по време на работа в световен мащаб. Всъщност, болката в гърба е приписана като втората най-често срещана причина за посещенията в лекарски кабинети, превишаващи броя им само чрез инфекции на горните дихателни пътища. Приблизително 80 процента от населението ще преживее някакъв вид болка в гърба поне веднъж през целия си живот. Гърбът е сложна структура, съставена от кости, стави, връзки и мускули, сред другите меки тъкани. Поради това, наранявания и / или влошени условия, като например херния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Болка тревожност депресияВсеки е изпитвал болка, но има и такива с депресия, тревожност или и двете. Комбинирайте това с болка и тя може да стане доста интензивна и трудна за лечение. Хората, които страдат от депресия, тревожност или и двете са склонни да изпитват силна и дългосрочна болка повече, отколкото другите хора.
Начинът безпокойство, депресията и болката се припокриват една с друга се наблюдава при хронични и при някои инвалидизиращи болкови синдроми, т.е. болка в кръста, главоболие, нервна болка и фибромиалгия. Психичните нарушения допринасят за интензивността на болката и също така увеличават риска от инвалидност.
депресия: A (голямо депресивно разстройство или клинична депресия) е често срещано, но сериозно разстройство на настроението. Той причинява тежки симптоми, които влияят върху това как се чувства, мисли и как се справя с ежедневните дейности, т.е. сън, хранене и работа. За да бъде диагностицирана депресия, симптомите трябва да присъстват поне две седмици.
Постоянно тъжно, тревожно или „празно“ настроение.
Чувство на безнадеждност, песимистично.
Раздразнителност.
Чувство за вина, безполезност или безпомощност.
Загуба на интерес или удоволствие от дейности.
Намалена енергия или умора.
Преместване или говорене бавно.
Чувствате се неспокойни и имате проблеми със седенето неподвижно.
Трудност съсредоточаване, помняне или вземане на решения.
Трудности със съня, събуждане рано сутрин и недоспане.
Промени в апетита и теглото.
Мисли за смърт или самоубийство и опити за самоубийство.
Болки или болки, главоболия, спазми или проблеми с храносмилането без ясна физическа причина и / или които не улесняват лечението.
Не всеки, който е депресиран, изпитва всеки симптом. Някои изпитват само няколко симптома, докато други могат да изпитат няколко. Няколко постоянни симптома в допълнение към лошото настроение са длъжен за диагностициране на тежка депресия. Тежестта и честотата на симптомите заедно с продължителността ще варират в зависимост от индивида и неговото конкретно заболяване. Симптомите също могат да варират в зависимост от стадия на заболяването.
ДЕПРЕСИЯ НА АНАЛИЗА НА БОЛЕСТИТЕ
Цели:
Каква е връзката?
Каква е неврофизиологията зад него?
Какви са основните последици?
Мозъчните промени в болката
Фигура 1 Мозъчни пътеки, региони и мрежи, участващи в остра и хронична болка
Davis, KD et al. (2017) Мозъчни образни тестове за хронична болка: медицински, правни и етични въпроси и препоръки Nat. Rev. Neurol. Дой: 10.1038 / nrneurol.2017.122
БОЛЕСТИ, ЗАБОЛЯВАНИЯ И ДЕПРЕСИЯ
Заключение:
Болката, особено хроничната, е свързана с депресия и безпокойство
Физиологичните механизми, водещи до тревожност и депресия, могат да имат многофакторен характер
Болката причинява промени в мозък структура и функция
Тази промяна в структурата и функцията може да промени способността на мозъка да модулира болката, както и настроението за контрол.
Когато сензорната система е засегната от нараняване или заболяване, нервите в тази система не могат да работят правилно, за да предават усещания и чувства в мозъка. Това често допринася за чувство на изтръпване или липса на усещане. Въпреки това, в определени случаи, когато тази система е повредена, хората могат да изпитат болка в засегнатата област.
Невропатична болка не започва внезапно и не преминава бързо; това е хронична болка състояние, което води до постоянни симптоми на болка. За повечето хора интензивността на техните симптоми може да нараства и да намалява през целия ден. Въпреки че се предполага, че невропатичната болка е свързана със здравословни проблеми на периферните нерви, като невропатия, причинена от диабет или спинална стеноза, нараняванията на мозъка или гръбначния мозък могат също да доведат до хронична невропатична болка. Невропатичната болка се нарича още нервна болка.
Невропатичната болка може да бъде противопоставена на ноцицептивната болка. Невропатичната болка не се развива поради някакво конкретно обстоятелство или външен стимул, а по-скоро симптомите се появяват просто защото нервната система може да не работи съответно. Всъщност хората също могат да изпитват невропатична болка, дори когато болната или наранена част от тялото всъщност не е там. Това състояние се нарича фантомна болка в крайниците, която може да се появи при хора след ампутация.
Ноцицептивната болка обикновено е остра и се развива в отговор на специфично обстоятелство, като например когато някой получи внезапно нараняване, като удряне на пръст с чук или убождане на пръст при ходене бос. Освен това, ноцицептивната болка има тенденция да изчезне, след като засегнатото място заздравее. Тялото съдържа специализирани нервни клетки, известни като ноцицептори, които откриват вредни стимули, които могат да увредят тялото, като екстремна топлина или студ, натиск, прищипване и излагане на химикали. След това тези предупредителни сигнали се предават по нервната система към мозъка, което води до ноцицептивна болка.
Какви са рисковите фактори за невропатична болка?
Всичко, което допринася за липса на функция в рамките на сензорната нервна система, може да доведе до невропатична болка. Като такива, проблемите със здравето на нервите от синдрома на карпалния тунел или подобни състояния могат в крайна сметка да предизвикат невропатична болка. Травмата, водеща до нараняване на нерва, може да доведе до невропатична болка. Други състояния, които биха могли да предразположат хората към развитие на невропатична болка, включват: диабет, дефицит на витамини, рак, ХИВ, инсулт, множествена склероза, херпес зостер и дори някои лечения на рак.
Какви са причините за невропатичната болка?
Има много причини, поради които хората могат да развият невропатична болка. Но на клетъчно ниво едно от обясненията е повишеното освобождаване на определени рецептори, които показват, че болката, заедно с намалената способност на нервите да модулират тези сигнали, води до усещане за болка, произхождаща от засегнатата област. Освен това, в гръбначния мозък, областта, която проявява болезнени признаци, се пренарежда със съответните промени в хормоните и загуба на нормално функциониращи подвижни тела. Тези промени водят до усещане за болка при липса на външна стимулация. В мозъка способността за блокиране на болката може да бъде засегната след нараняване, като инсулт или травма от нараняване. С течение на времето се случват допълнителни увреждания на клетките и усещането за болка продължава. Невропатичната болка също е свързана с диабет, хроничен прием на алкохол, някои видове рак, дефицит на витамин В, заболявания, други заболявания, свързани с нервите, токсини и специфични лекарства.
Какви са симптомите на невропатичната болка?
За разлика от други неврологични състояния, идентифицирането на невропатичната болка може да бъде предизвикателство. Въпреки това може да има няколко, ако има такива, обективни сигнала. Здравните специалисти трябва да дешифрират и превеждат набор от думи, които пациентите използват, за да опишат болката си. Пациентите могат да опишат симптомите си като остри, тъпи, горещи, студени, чувствителни, сърбящи, дълбоки, парещи, парещи, сред множество други описателни термини. Освен това някои пациенти могат да изпитат болка при леко докосване или натиск.
В опит да се помогне да се определи колко болка могат да изпитат пациентите, често се използват различни скали. От пациентите се изисква да оценят болката си според визуална скала или цифрова графика. Съществуват много примери за скали за болка, като тази, показана по-долу. Често снимките на лица, изобразяващи различни нива на болка, могат да бъдат полезни, когато хората имат трудности да опишат количеството болка, което изпитват.
Хронична болка и психично здраве
За мнозина въздействието на хроничната болка може да не се ограничава до болката; може също така да повлияе негативно на психическото им състояние. Нови изследователски проучвания, проведени от учени от Северозападния университет в Чикаго, могат да обяснят защо хората, които имат хронична болка, страдат и от привидно несвързани здравословни проблеми, като депресия, стрес, липса на сън и затруднено концентриране.
Оценката показа, че хората с хронична болка показват различни региони на мозъка, които винаги са активни, по-специално областта, свързана с настроение и внимание. Това непрекъснато действие пренарежда нервните връзки от мозъка и оставя страдащите от хронична болка изложени на по-голям риск от психологически проблеми. Изследователите предполагат, че постоянното получаване на сигнали за болка може да доведе до умствено пренастройване, което се отразява неблагоприятно на ума. Пренасочването принуждава мозъците им да отделят умствени ресурси по различен начин, за да се справят с ежедневните задачи, от математика, през припомняне на списък за пазаруване, до чувство на щастие.
Връзката между болката и мозъка е добре записана, поне анекдотично, и много здравни специалисти казват, че са видели от първа ръка как психическото състояние на пациента може да се понижи, когато понася хронична болка. Погрешните схващания за връзката болка-мозък може да са се появили от липсата на доказателства, че болката има измеримо, трайно влияние върху мозъка. Изследователите очакват, че с допълнителни изследвания на механизмите за това как хроничната болка прави хората по-податливи на разстройства на настроението, хората ще имат способността да управляват по-добре цялостното си благосъстояние.
Култура и хронична болка
Много неща допринасят за начина, по който изпитваме и изразяваме болката, но наскоро беше предложено от изследователите, че културата има пряка връзка с изразяването на болката. Нашето възпитание и обществени ценности влияят върху начина, по който изразяваме болката, както и нейната собствена природа, интензивност и продължителност. Тези променливи обаче не са толкова очевидни, колкото социално-психологическите стойности, като възраст и пол.
Изследванията сочат, че хроничната болка е многостранен процес и едновременното взаимодействие между патофизиологията, когнитивните, афективните, поведенческите и социокултурните фактори се обобщава до това, което се нарича преживяване на хронична болка. Оказа се, че хроничната болка се изпитва по различен начин сред пациентите от различни култури и етноси.
Някои култури насърчават изразяването на болка, особено в южното Средиземноморие и Близкия изток. Други хора го потискат, както в многото уроци за нашите деца как да се държим смело и да не плачем. Болката се признава като част от човешкия опит. Склонни сме да приемем, че комуникацията за болката ще премине безпроблемно културните граници. Но хората, страдащи от болка, са подчинени на маниерите, които техните цивилизации са ги обучили да изпитват и изразяват болка.
Както лицата, страдащи от болка, така и здравните специалисти изпитват трудности при общуването на болката през етническите граници. Във въпрос като болката, където ефективната комуникация може да има далечни последици за медицинските грижи, качеството на живот и потенциалното оцеляване, ролята на културата в общуването на болката остава подценена. Постоянната болка е многоизмерна, съставна среща, образувана от преплитане и съвместно влияние на биологични и психосоциални фактори. Познаването на кулминацията на тези фактори е от решаващо значение за разбирането на разликите в нейното проявление и управление.
Как се диагностицира невропатичната болка?
Диагнозата на невропатичната болка се основава на допълнителна оценка на анамнезата на индивида. Ако се подозира основно увреждане на нервите, тогава анализът на нервите заедно с тестването може да бъде оправдан. Най-често срещаното средство за преценка дали даден нерв е увреден или не е използването на електродиагностично лекарство. Тази медицинска подспециалност използва техники за изследване на нервната проводимост с електромиелография (NCS/EMG). Клиничната оценка може да покаже доказателство за загуба на работа и може да включва оценка на леко докосване, способност за разграничаване на остра от тъпа болка и способност за разпознаване на температурата, както и оценка на вибрациите.
След завършване на задълбочен клиничен преглед може да се планира електродиагностичният анализ. Тези изследвания се провеждат от специално обучени невролози и физиатри. Ако се подозира невропатия, трябва да се извърши търсене на обратими причини. Това може да включва кръвна функция за дефицит на витамини или проблеми с щитовидната жлеза и образни изследвания за изключване на структурна лезия, засягаща гръбначния мозък. В зависимост от резултатите от това изследване може да има средства за намаляване на интензивността на невропатията и евентуално намаляване на болката, която пациентът изпитва.
За съжаление, при много състояния дори добрият контрол на основната причина за невропатията не може да обърне невропатичната болка. Това често се наблюдава при пациенти с диабетна невропатия. В редки случаи може да има признаци на промени в кожата и модела на растеж на косата в засегнатия регион. Тези промени могат да бъдат свързани с промени в изпотяването. Ако са налице, тези промени могат да помогнат за идентифициране на вероятното присъствие на невропатична болка, свързана със състояние, известно като комплексен регионален болков синдром.
Прозрение на д-р Алекс Химензе
Невропатичната болка е състояние на хронична болка, което обикновено се свързва с директно увреждане или нараняване на нервната система или нервите. Този тип болка е различен от ноцицептивната болка или типичното усещане за болка. Ноцицептивната болка е остро или внезапно усещане за болка, което кара нервната система да изпраща сигнали за болка веднага след настъпването на травмата. При невропатична болка обаче пациентите могат да изпитат прострелваща, пареща болка без директни увреждания или наранявания. Разбирането на възможните причини за невропатичната болка на пациента спрямо всеки друг вид болка може да помогне на здравните специалисти да намерят по-добри начини за лечение на състояния на хронична болка.
Какво е лечението на невропатичната болка?
Използват се различни лекарства за лечение на невропатична болка. Повечето от тези лекарства се използват извън етикета, което означава, че лекарството е одобрено от FDA за лечение на различни състояния и след това е признато за благоприятно за лечение на невропатична болка. Трицикличните антидепресанти, като амитриптилин, нортриптилин и дезипрамин, се предписват за лечение на невропатична болка от няколко години.
Някои хора смятат, че те могат да бъдат много ефективни за облекчение. Доказано е, че други видове антидепресанти предлагат известно облекчение. При някои пациенти се използват селективни инхибитори на обратното захващане на серотонин или SSRI, като пароксетин и циталопрам, и други антидепресанти, като венлафаксин и бупропион. Друго често лечение на невропатична болка включва лекарства против припадъци, включително карбамазепин, фенитоин, габапентин, ламотригин и други.
При остри случаи на болезнена невропатия, които не се повлияват от брокери от първа линия, лекарствата, които обикновено се използват за лечение на сърдечни аритмии, могат да бъдат от полза; те обаче могат да доведат до значителни странични ефекти и често трябва да се наблюдават внимателно. Лекарствата, прилагани директно върху кожата, могат да предложат скромни до осезаеми ползи за някои пациенти. Често използваните форми включват лидокаин (под формата на пластир или гел) или капсаицин.
Лечението на невропатичната болка зависи от основната причина. Ако причината е обратима, тогава периферните нерви могат да се регенерират и болката ще отшуми; въпреки това, това намаляване на болката може да отнеме няколко месеца до години. Няколко други алтернативни възможности за лечение, включително хиропрактика и физическа терапия, също могат да бъдат използвани, за да се помогне за облекчаване на напрежението и стреса покрай нервите, като в крайна сметка помага за подобряване на болезнените симптоми.
Каква е прогнозата за невропатична болка?
Много хора с невропатична болка са в състояние да получат някаква помощ, дори когато болката им продължава. Въпреки че невропатичната болка не е опасна за пациента, наличието на хронична болка може да повлияе негативно на качеството на живот. Пациентите с хронична нервна болка могат да страдат от лишаване от сън или нарушения на настроението, включително депресия, тревожност и стрес, както беше споменато по-горе. Поради присъщата алопеция и липсата на сензорна обратна връзка, пациентите са изложени на риск от развитие на нараняване или инфекция или несъзнателно причиняване на ескалация на настоящо нараняване. Ето защо е от съществено значение да потърсите незабавна медицинска помощ и да следвате специфични насоки, насочени от медицински специалист за безопасност и предпазливост.
Може ли невропатичната болка да бъде предотвратена?
Най-добрият начин за предотвратяване на невропатичната болка е да се избегне развитието или прогресията на невропатията. Мониторинг и промяна на начина на живот, включително ограничаване на употребата на алкохол и тютюн; поддържане на здравословно тегло за намаляване на риска от диабет, дегенеративно ставно заболяване или инсулт; и наличието на страхотна ергономична форма по време на работа или при практикуване на хобита за намаляване на риска от повтарящи се стресови наранявания са стратегии за намаляване на вероятността от развитие на невропатия и потенциална невропатична болка. Не забравяйте да потърсите незабавна медицинска помощ в случай на симптоми, свързани с невропатична болка, за да продължите с най-подходящия подход за лечение. Обхватът на нашата информация е ограничен до хиропрактика, както и до наранявания и състояния на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителни теми: Болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати дни по време на работа в световен мащаб. Всъщност, болката в гърба е приписана като втората най-често срещана причина за посещенията в лекарски кабинети, превишаващи броя им само чрез инфекции на горните дихателни пътища. Приблизително 80 процента от населението ще преживее някакъв вид болка в гърба поне веднъж през целия си живот. Гърбът е сложна структура, съставена от кости, стави, връзки и мускули, сред другите меки тъкани. Поради това, наранявания и / или влошени условия, като например херния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Ако сензорната система бъде засегната от нараняване или заболяване, нервите в тази система не могат да функционират при предаването на усещане към мозъка. Това може да доведе до усещане за изтръпване или липса на усещане. В някои случаи, когато сензорната система е наранена, хората могат да изпитат болка в засегнатата област. Невропатичната болка не започва бързо или завършва бързо. Това е хронично състояние, което води до симптоми на постоянна болка. За мнозина интензивността на симптомите може да идва и да си отива през целия ден. Смята се, че невропатичната болка е свързана с проблеми с периферните нерви, т.е. невропатия, причинена от диабет, спинална стеноза, нараняване на мозъка или гръбначния мозък също може да доведе до хронична невропатична болка.
НЕВРОПАТИЧНА БОЛКА
Цели:
Какво е това?
Каква е патофизиологията зад това?
Какви са причините?
Кои са някои от пътищата
Как можем да го поправим?
НЕВРОПАТИЧНА БОЛКА
Болка, инициирана или причинена от първична лезия или дисфункция в соматосензорната нервна система.
Невропатичната болка обикновено е хронична, трудна за лечение и често устойчива на стандартно аналгетично лечение.
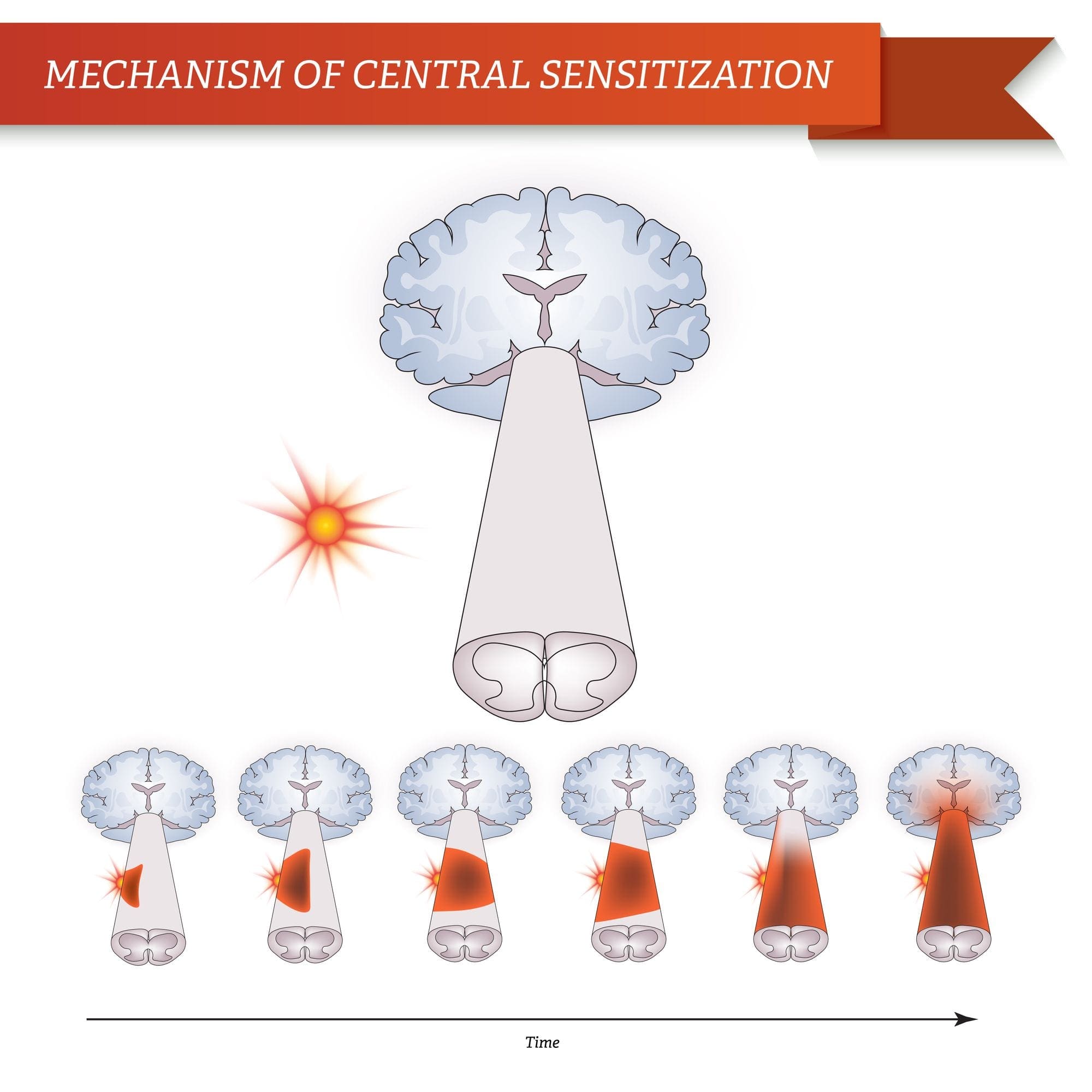
ПАТОГЕНЕЗА НА НЕВРОПАТИЧНА БОЛКА
ПЕРИФЕРНИ МЕХАНИЗМИ
След лезия на периферните нерви невроните стават по-чувствителни и развиват необичайна възбудимост и повишена чувствителност към стимулация
Това е известно като... периферна сенсибилизация!
ЦЕНТРАЛНИ МЕХАНИЗМИ
В резултат на продължаваща спонтанна активност, възникваща в периферията, невроните развиват повишена фонова активност, разширени рецептивни полета и повишени отговори на аферентни импулси, включително нормални тактилни стимули
Това е известно като…Централна сенсибилизация!
ЧЕСТИ ПРИЧИНИ
Лезии или заболявания на соматосензорната нервна система могат да доведат до променено и нарушено предаване на сензорни сигнали в гръбначния мозък и мозъка; честите състояния, свързани с невропатична болка, включват:
Postherpetic neuralgia
Тригеминална невралгия
Болезнена радикулопатия
Диабетна невропатия
HIV инфекция
проказа
ампутиране
Болка при нараняване на периферните нерви
Инсулт (под формата на централна болка след инсулт)
ФАНТОМНА БОЛКА В КРАЙНИКАТА И РАЗОШЕНА РЕАЛНОСТ
Фантомна болка в крайниците и AR
НЕВРОГЕННО ВЪЗПАЛЕНИЕ
Цели:
Какво е това?
Каква е патофизиологията зад това?
Какви са причините?
Как можем да го поправим?
НЕВРОГЕННО ВЪЗПАЛЕНИЕ
Неврогенно възпаление е невронно предизвикан локален възпалителен отговор, характеризиращ се с вазодилатация, повишена съдова пропускливост, дегранулация на мастоцитите и освобождаване на невропептиди, включително SP и пептид, свързан с калцитонин ген (CGRP)
Изглежда, че играе важна роля в патогенезата на множество заболявания, включително мигрена, псориазис, астма, фибромиалгия, екзема, розацея, дистония и множествена химическа чувствителност
ЧЕСТИ ПРИЧИНИ
Има множество пътища, по които може да се инициира неврогенно възпаление. Добре е документирано, използвайки както животински модели, така и изолирани неврони in vitro, че капсаицинът, топлината, протоните, брадикининът и триптазата са регулатори нагоре по веригата на вътреклетъчния приток на калций, което води до освобождаване на възпалителен невропептид. Обратно, смята се, че простагландините E2 и I2, цитокини, интерлевкин-1, интерлевкин-6 и тумор некрозис фактор не предизвикват самостоятелно освобождаване на невротрансмитери, а по-скоро възбуждат сензорните неврони и по този начин понижават прага за активиране и предизвикват увеличено освобождаване на невропептиди.
Докато неврогенното възпаление е широко проучено и добре документирано в периферните тъкани, доскоро концепцията за неврогенно възпаление в ЦНС остава до голяма степен неизследвана. Като се има предвид способността на неврогенното възпаление да повлияе на съдовата пропускливост и да доведе до генезис на оток, сега е широко изследван потенциалът му да повлияе на пропускливостта на ВВВ и вазогенния оток в мозъка и гръбначния мозък при различни патологични състояния.
Всички ни бяха учили като деца, че има 5 сетива: зрение, вкус, звук, мирис и докосване. Първоначалните четири сетива използват ясни, отделни органи, като очи, вкусови рецептори, уши и нос, но как точно сетивното усещане на тялото се докосва? Докосването се усеща върху цялото тяло, както отвътре, така и отвън. Няма отделен орган, който да е отговорен за усещането за докосване. По-скоро около цялото тяло има малки рецептори или нервни окончания, които усещат докосването там, където се случва, и изпраща сигнали до мозъка с информация относно вида на докосването, което се е случило. Тъй като вкусовата пъпка на езика разпознава вкуса, механорецепторите са жлези в кожата и на други органи, които откриват усещанията за допир. Те са известни като Механорецептори защото са проектирани да откриват механични усещания или разлики в налягането.
Роля на механорецепторите
Човек разбира, че е изпитал усещане, след като органът, отговорен за откриването на това специфично сетиво, изпрати съобщение до мозъка, който е основният орган, който обработва и подрежда цялата информация. Съобщенията се изпращат от всички области на тялото към мозъка чрез проводници, наречени неврони. Има хиляди малки неврони, които се разклоняват към всички области на човешкото тяло, а на краищата на много от тези неврони са механорецептори. За да демонстрираме какво се случва, когато докоснете обект, ще използваме пример.
Представете си комар да кацне на ръката ви. Щамът на това насекомо, толкова лек, стимулира механорецепторите в тази конкретна област на ръката. Тези механорецептори изпращат съобщение по неврона, с който са свързани. Невронът се свързва чак до мозъка, който получава съобщението, че нещо докосва тялото ви на точното място на конкретния механорецептор, изпратил съобщението. Мозъкът ще действа с този съвет. Може би ще каже на очите да погледнат областта на ръката, която е открила подписа. И когато очите казват на мозъка, че има комар на ръката, мозъкът може да каже на ръката бързо да го махне. Така работят механорецепторите. Целта на статията по-долу е да демонстрира, както и да обсъди подробно функционалната организация и молекулярните детерминанти на механорецепторите.
Усещане за докосване: Функционална организация и молекулярни детерминанти на механочувствителните рецептори
абстрактен
Кожните механорецептори са локализирани в различните слоеве на кожата, където откриват широк спектър от механични стимули, включително лека четка, разтягане, вибрации и вреден натиск. Това разнообразие от стимули е съчетано от разнообразен набор от специализирани механорецептори, които реагират на кожна деформация по специфичен начин и предават тези стимули към по-високи мозъчни структури. Изследванията върху механорецепторите и генетично поносимите сетивни нервни окончания започват да разкриват механизми на докосване. Работата в тази област предостави на изследователите по-задълбочено разбиране на организацията на веригата, лежаща в основата на възприятието за докосване. Нови йонни канали се появиха като кандидати за трансдукционни молекули и свойствата на механично затворени токове подобриха нашето разбиране за механизмите на адаптация към тактилни стимули. Този преглед подчертава напредъка, постигнат в характеризирането на функционалните свойства на механорецепторите в космата и гола кожа и йонните канали, които откриват механични входове и адаптация на механорецепторите на формата.
Докосването е откриване на механичен стимул, въздействащ върху кожата, включително безобидни и вредни механични стимули. Това е основен смисъл за оцеляването и развитието на бозайниците и човека. Контактът на твърди предмети и течности с кожата дава необходимата информация на централната нервна система, която позволява изследване и разпознаване на околната среда и инициира движение или планирано движение на ръката. Докосването също е много важно за чиракуването, социалните контакти и сексуалността. Усещането за допир е най-слабо уязвимото, въпреки че може да бъде изкривено (хиперестезия, хипестезия) при много патологични състояния.1-3
Реакциите на докосване включват много прецизно кодиране на механична информация. Кожните механорецептори са локализирани в различните слоеве на кожата, където откриват широк спектър от механични стимули, включително лека четка, разтягане, вибрации, отклоняване на косата и вреден натиск. Това разнообразие от стимули е съчетано от разнообразен набор от специализирани механорецептори, които реагират на кожна деформация по специфичен начин и предават тези стимули към по-високи мозъчни структури. Соматосензорните неврони на кожата попадат в две групи: нископрагови механорецептори (LTMR), които реагират на доброкачествено налягане и високопрагови механорецептори (HTMR), които реагират на вредно механично стимулиране. LTMR и HTMR клетъчните тела се намират в ганглиите на дорзалния корен (DRG) и черепните сензорни ганглии (тригеминални ганглии). Нервните влакна, свързани с LTMR и HTMR, се класифицират като Ap-, Ap- или C-влакна въз основа на скоростите на проводимост на техния потенциал на действие. C влакната са немиелинизирани и имат най-бавни скорости на проводимост (~2 m/s), докато A? и А? влакната са леко и силно миелинизирани, като показват междинни (~12 m/s) и бързи (~20 m/s) скорости на проводимост, съответно. LTMR също се класифицират като бавно или бързо адаптиращи се отговори (SA- и RA-LTMR) според скоростта им на адаптация към продължителен механичен стимул. Освен това те се отличават с кожните крайни органи, които инервират и предпочитаните от тях стимули.
Способността на механорецепторите да откриват механични сигнали разчита на наличието на йонни канали на механотрансдюсер, които бързо трансформират механичните сили в електрически сигнали и деполяризират възприемчивото поле. Тази локална деполяризация, наречена рецепторен потенциал, може да генерира потенциали на действие, които се разпространяват към централната нервна система. Въпреки това, свойствата на молекулите, които медиират механотрансдукцията и адаптацията към механичните сили, остават неясни.
В този преглед ние предоставяме преглед на свойствата на механорецепторите на бозайници при безвредно и вредно докосване в окосмена и гола кожа. Ние също така разглеждаме последните познания за свойствата на механично затворените токове в опит да обясним механизма на адаптация на механорецептора. И накрая, ние правим преглед на скорошния напредък, постигнат в идентифицирането на йонни канали и свързаните с тях протеини, отговорни за генерирането на механизирани токове.
Безобидно докосване
LTMR, свързани с космения фоликул
Космените фоликули представляват мини-органи, произвеждащи косми, които откриват леко докосване. Влакната, свързани с космените фоликули, реагират на движението на косъма и неговата посока чрез изстрелване на влакове от потенциали на действие при появата и отстраняването на стимула. Те са бързо адаптиращи се рецептори.
Котка и заек. При котешка и заешка козина космените фоликули могат да бъдат разделени на три вида космени фоликули, косми на пух, косми на Guard и Tylotrichs. Пуховите косми (подкосъм, вълна, велус)4 са най-многобройните, най-късите и фини косми на козината. Те са вълнообразни, безцветни и се появяват на групи от по два до четири косъма от общ отвор в кожата. Гвардейските косми (монотрихи, overhears, tophair)4 са леко извити, пигментирани или непигментирани, и излизат поотделно от устията на техните фоликули. Тилотрихите са най-малко многобройни, най-дългите и най-дебели косми.5,6 Те са пигментирани или непигментирани, понякога и двете и излизат поотделно от фоликул, който е заобиколен от бримка от капилярни кръвоносни съдове. Сетивните влакна, които доставят на космения фоликул, се намират под мастната жлеза и се приписват на A? или A?-LTMR влакна.7
В близост до долната част на косъма, точно под нивото на мастната жлеза е пръстенът от копиевидни пило-Ruffini окончания. Тези сензорни нервни окончания са разположени в спираловиден ход около косъма в рамките на съединителната тъкан, образуваща космения фоликул. В космения фоликул има и свободни нервни окончания, някои от които образуват механорецептори. Често допирните клетки (виж гола кожа) заобикалят областта на шията на тилотриховия фоликул.
Свойствата на миелинизираните нервни окончания в окосмена кожа на котки и зайци са изследвани интензивно през периода 1930–1970 г. (преглед в Hamann, 1995).8 Забележително е, че Браун и Иго, изучавайки 772 единици с миелинизирани аферентни нервни влакна в котешките нерви и заек, имат класифицирани реакции в три типа рецептори, съответстващи на движенията на космите надолу (рецептори тип D), Guard коса (тип G рецептори) и Tylotrich коса (тип T рецептор).9 Всички отговори на аферентните нервни влакна са събрани в бързо адаптиращия се рецептор от тип I (RA I) чрез противопоставяне на рецептора на Пачини, наречен RA II. Механорецепторите на RA I откриват скоростта на механичния стимул и имат остра граница. Те не откриват термични вариации. Burgess et al. също така описва бързо адаптиращ се полеви рецептор, който реагира оптимално на поглаждане на кожата или движение на няколко косми, което се приписва на стимулиране на пило-Руфини окончания. Нито един от отговорите на космените фоликули не се дължи на активността на C влакното.10
Мишки. В гръбната окосмена кожа на мишките са описани три основни типа космени фоликули: зигзаг (около 72%), шило/аушен (около 23%) и предпазител или тилотрих (около 5%).11-14 Зигзаг и шило/ Auchenne космените фоликули произвеждат по-тънките и къси косми и са свързани с една мастна жлеза. Защитните или тилотриховите косми са най-дългите от видовете космени фоликули. Те се характеризират с голяма космена луковица, свързана с две мастни жлези. Защитните косми и космите от шило/аушен са подредени в итеративен, равномерно разположен модел, докато зигзагообразните косми гъсто заселват кожните области около двата по-големи типа космени фоликули [Фиг. 1 (A1, A2 и A3)].
Фигура 1. Организация и проекции на кожни механорецептори. При окосмена кожа леката четка и докосването се засичат главно от инервацията около космените фоликули: шило/ошен (A1), зигзаг (A2) и предпазител (A3). Космите от шило/ашена са тройно инервирани от C-LTMR ланцетни окончания (A4), A?-LTMR и A? бързо адаптиращ се-LTMR (A6). Зигзагообразните космени фоликули са по-късите косми и се инервират както от C-LTMR (A4), така и от A? -LTMR ланцетни окончания (A5). Най-дългите предпазни космени фоликули се инервират от A? бързо адаптиращи се-LTMR надлъжни ланцетни краища (A6) и са свързани с A? бавно адаптиране-LTMR на края на сензорния купол (A7). Централните издатини на всички тези влакна завършват в отделни, но частично припокриващи се ламини на гръбначния рог на гръбначния мозък (C-LTMR в ламина II, Ap-LTMR в ламина III и Ap-LTMR в ламина IV и V). Проекциите на LTMR, които инервират същите или съседни космени фоликули, са подравнени, за да образуват тясна колона в гръбначния рог на гръбначния мозък (B1 в сиво). Само при окосмена кожа субпопулация от свободни крайници на С-влакна инервира епидермиса и реагира на приятно докосване (A8). Тези C-touch влакна не реагират на вредно докосване и тяхното движение все още не е известно (B2). При гола кожа безобидното докосване се медиира от четири типа LTMR. Клетъчно-невритният комплекс на Меркел е в базалния слой на епидермиса (С1). Този механорецептор се състои от подреждане между много клетки на Меркел и разширен нервен терминал от един A? фибри. Клетките на Меркел показват пръстовидни процеси, контактуващи с кератиноцитите (С2). Окончанието на Ruffini е локализирано в дермата. Това е тънък капсулиран сензорен край с форма на пура, свързан с A? влакно (С3). Корпускулата на Майснер, свързана с A? нервно окончание и се намира в дермалните папили. Този капсулиран механорецептор се състои от опаковани поддържащи клетки, подредени като хоризонтални ламели, заобиколени от съединителна тъкан (C4). Пацинова телце е по-дълбокият механорецептор. Едно единствено А? немиелинизираното нервно окончание завършва в центъра на това голямо яйцевидно телце, изградено от концентрични ламели. Проекциите на тези Ap-LTMR влакна в гръбначния мозък са разделени на два клона. Главният централен клон (B3) се издига в гръбначния мозък в ипсилатералната дорзална част, образувайки клиновидни или грацилни пучки (B5) на ниво медула, където първичните аференти правят първия си синапс (B6). Вторичните неврони правят сензорна декусация (B7), за да образуват тракт върху медиалния лемнискус, който се издига през мозъчния ствол към средния мозък, по-специално в таламуса. Вторичният клон на LTMR завършва в дорзалния рог в ламина II, IV, V и пречи на предаването на болката (B4). Вредното докосване се открива от свободните нервни окончания в епидермиса както на окосмена (A9), така и на гола кожа (C7). Тези механорецептори са краят на Ap-HTMR и C-HTMR в близък контакт със съседни кератиноцити (C6). Ap-hTMR завършва в ламината I и V; C-HTMR завършва в ламината I и II (B8). На нивото на дорзалния рог на гръбначния мозък първичните аферентни HTMR правят синапси с вторични неврони, които пресичат средната линия и се изкачват до по-високата мозъчна структура в преднолатералния фасцикул (B9, B10).
Наскоро Джинти и сътрудниците му използваха комбинация от молекулярно-генетично маркиране и подходи за соматотопно ретроградно проследяване, за да визуализират организацията на периферните и централните аксонални окончания на LTMR при мишки.15 Техните открития подкрепят модел, в който индивидуалните характеристики на сложен тактилен стимул са се извлича от трите типа космени фоликули и се предава чрез дейностите на уникални комбинации от A?-, A?- и C- влакна до гръбния рог.
Те показаха, че генетичното маркиране на тирозин хидроксилаза положителни (TH+) DRG неврони характеризира популация от непептидергични сензорни неврони с малък диаметър и позволява визуализация на C-LTMR периферни окончания в кожата. Изненадващо е установено, че аксоналните клони на отделни C-LTMR се образуват и образуват надлъжни ланцетни краища, които са тясно свързани със зигзаг (80% от краищата) и шило/аушен (20% от краищата), но не и тилотрични космени фоликули [Фиг. 1 (A4)]. Дълго време се смяташе, че надлъжните ланцетни краища принадлежат изключително на A?-LTMR и затова беше неочаквано, че краищата на C-LTMR ще образуват надлъжни ланцетни окончания.15 Тези C-LTMR имат междинна адаптация в сравнение с бавно и бързо адаптиращите се миелинизирани механорецептори [фиг. 2 (C1)].
Фигура 2. Тактилни рецептори при бозайници: Кожните тактилни рецептори се диференцират в безвредно докосване, поддържано от множество рецептори с нисък механичен праг (LTMR) в гола и окосмена кожа и вредно докосване, поддържано от рецептор с висок механичен праг (HTMR). Те образуват свободни от нерви окончания, които завършват главно в епидермиса. (A) Гладка кожа. A1: Мейснеровите телца откриват движение на кожата и подхлъзване на обект в ръката. Те са важни за подаване на предмет и сръчност. Рецепторите бързо се адаптират към стимула, свързани са с A? влакна и рядко до C влакна и имат голямо рецепторно поле. A2: Телцата на Руфини откриват разтягане на кожата и са важни за откриване на позицията на пръста и подавания предмет. Рецепторът бавно се адаптира към стимула и поддържа активност, докато се прилага стимулът. Рецепторите са свързани към A? влакна и имат голямо рецептивно поле. A3: Пачинови телца са по-дълбоко в дермата и откриват вибрации. Рецепторите са свързани към A? влакна; те бързо се адаптират към стимула и имат най-голямо рецептивно поле. (B) Цяла кожа. B1: Меркел-клетъчните комплекси присъстват както в голата кожа, така и около косата. Те са плътно изразени в ръката и са важни за възприемането на текстурата и най-фината дискриминация между две точки. Те са отговорни за прецизността на пръстите. Рецепторите са свързани към A? влакна; те бавно се адаптират към стимула и имат кратко рецептивно поле. B2: Вредни при докосване HTMR с много бавна адаптация към стимула, т.е. активни, докато се прилага ноцицептивният стимул. Те се образуват от свободния нервен край на A? и С-влакна, свързани с кератиноцитите. (C) Окосмена кожа. C1: Космите фоликули са свързани с различните типове коса. При мишки гвардейските косми са по-дълги и слабо изразени, шилото/ашената са със среден размер, а зигзагообразните са най-малките и най-гъсто изразени косми. Те са свързани с А? влакна, но и към A? и C-LTMRs влакна за шило/ошен и зизаг коса. Те засичат движението на косата, включително приятно докосване по време на ласка. Те се адаптират бързо или със средна кинетика към стимула. C2: C-touch нервните окончания съответстват на подтип на края на C влакната със свободен край, характеризиращ се с нисък механичен праг. Те трябва да кодират приятно усещане, предизвикано от ласка. Те умерено се адаптират към стимули и имат кратко рецептивно поле. Предполагаемите механочувствителни (MS) йонни канали, експресирани в различните тактилни рецептори, са посочени в съответствие с предварителните данни и обобщават настоящата хипотеза, която се оценява.
Втората основна идентифицирана популация се отнася до края на A?-LTMR в Awl/Auchenne и зигзагообразни фоликули, които да бъдат сравнени с Down космения фоликул, подробно изследван при котка и заек. Ginty и сътрудниците му показаха, че TrkB се експресира на високи нива в подгрупа от DRG неврони със среден диаметър. Вътреклетъчните записи, използващи ex vivo кожно-нервния препарат на белязани влакна, разкриват, че те проявяват физиологичните свойства на влакната, изследвани преди това при котки и зайци: изискана механична чувствителност (праг на фон Фрей < 0.07 mN), бързо адаптиране на реакции към надпрагови стимули, междинна проводимост скорости (5.8 ± 0.9 m/s) и тесни неизкривени сома шипове.15 Тези A?-LTMR образуват надлъжни ланцетни окончания, свързани с почти всеки зигзагообразен и шило/ашенен космен фоликул на ствола [Фиг. 1 (A5)].
И накрая, те показаха, че периферните окончания на бързо адаптиращия се A? LTMR образуват надлъжни ланцетни окончания, свързани с предпазни (или tylotrich) и шилови/ашенови космени фоликули [Фиг. 1 (A6)].15 Освен това космите на Guard са свързани и с клетъчен комплекс на Меркел, образуващ сензорен купол, свързан с A? бавно адаптиращ се LTMR [фиг. 1 (A7)].
В обобщение, почти всички зигзагообразни космени фоликули са инервирани както от C-LTMR, така и от A?-LTMR ланцетни окончания; космите от шило/аушен са трикратно инервирани от A? бързо адаптиращи се-LTMR, Ap-LTMR и C-LTMR ланцетни окончания; Защитните космени фоликули се инервират от A? бързо адаптиращи се-LTMR надлъжни ланцетни окончания и взаимодействат с A? бавно адаптиране-LTMR на края на сензорния купол. По този начин всеки миши космен фоликул получава уникални и инвариантни комбинации от LTMR окончания, съответстващи на неврофизиологично различни механосензорни крайни органи. Като се има предвид итеративното подреждане на тези три типа коса, Джинти и сътрудниците му предлагат окосмената кожа да се състои от итеративно повторение на периферна единица, съдържаща (1) една или две централно разположени предпазни косми, (2) ~20 околни косми от шило/аушен и (3 ) ~80 разпръснати зигзагообразни косми [фиг. 2 (C1)].
Проекция на гръбначния мозък. Централните проекции на A? бързо адаптиращите се LTMR, Ap-LTMR и C-LTMR завършват в отделни, но частично припокриващи се ламини (II, III, IV) на гръбначния рог на гръбначния мозък. В допълнение, централните терминали на LTMR, които инервират същите или съседни космени фоликули в рамките на периферна LTMR единица, са подравнени, за да образуват тясна LTMR колона в гръбначния рог на гръбначния мозък [фиг. 1 (B1)]. По този начин изглежда вероятно, че клин или колона от соматотопно организирани първични сензорни аферентни окончания в гръбния рог представлява подравняването на централните проекции на A?-, A?- и C-LTMR, които инервират една и съща периферна единица и откриват механични стимули, действащи върху една и съща малка група космени фоликули. Въз основа на броя на предпазните косми, шила/ашена и зигзагообразните косми на тялото и крайниците и броя на всеки LTMR подтип, Ginty и сътрудниците изчисляват, че гръбният рог на мишката съдържа 2,000 4,000 × 15 XNUMX LTMR колони, което съответства на приблизителния брой на LTMR LTMR единици.XNUMX
Освен това, аксоните от подтипове LTMR са тясно свързани един с друг, като имат преплетени издатини и преплетени ланцетни краища, които инервират един и същ космен фоликул. В допълнение, тъй като трите типа космени фоликули проявяват различни форми, размери и клетъчни състави, те вероятно имат различни отклоняващи или вибрационни настройващи свойства. Тези констатации са в съответствие с класическите неврофизиологични измервания при котка и заек, показващи, че A? RA-LTMR и A?-LTMR могат да бъдат диференциално активирани чрез отклоняване на различни типове космени фоликули.16,17
В заключение, докосването в окосмена кожа е комбинацията от: (1) относителните числа, уникални пространствени разпределения и различни морфологични и отклоняващи свойства на трите типа космени фоликули; (2) уникалните комбинации от окончания на подтип LTMR, свързани с всеки от трите типа космени фоликули; и (3) различни чувствителност, скорости на проводимост, модели на върхови влакове и адаптивни свойства на четирите основни класа свързани с космените фоликули LTMR, които позволяват на механосензорната система на космата кожа да извлича и предава на ЦНС сложните комбинации от качества, които определят докосване.
LTMR на свободни нервни окончания
По принцип свободните краища на C-влакна в кожата са HTMR, но субпопулация от C-влакна не реагира на вредно докосване. Тази подгрупа от тактилни C-влакна (CT) аференти представлява отделен тип немиелинизирани, нископрагови механорецептивни единици, съществуващи в космата, но не гола кожа на хора и бозайници [Фиг. 1 (A8)].18,19 CT обикновено се свързват с възприемането на приятна тактилна стимулация при телесен контакт.20,21
CT аферентите реагират на сили на вдлъбнатина в диапазона 0.3-2.5 mN и по този начин са толкова чувствителни към деформация на кожата, колкото много от A? аференти.19 По този начин характеристиките на адаптация на CT аферентите са междинни в сравнение с бавно и бързо адаптиращите се миелинизирани механорецептори. Рецептивните полета на човешки CT аференти са с приблизително кръгла или овална форма. Полето се състои от едно до девет малки отзивчиви петна, разпределени върху площ до 35 mm2.22 Хомоложните рецептори на мишката са организирани в модел от прекъснати петна, покриващи около 50% от площта в окосмената кожа [фиг. 60 (C2)].2
Доказателства от пациенти без миелинизирани тактилни аференти показват, че сигнализирането в CT влакната активира островната кора. Тъй като тази система е слаба в кодирането на дискриминационни аспекти на докосването, но е добре пригодена за кодиране на бавно, нежно докосване, CT влакната в окосмена кожа могат да бъдат част от система за обработка на приятни и социално значими аспекти на докосването.24 Активирането на CT влакна може също да имат роля в инхибирането на болката и наскоро беше предложено, че възпалението или травмата могат да променят усещането, предавано от C-влакна LTMR от приятно докосване към болка.25,26
По кой път пътуват CT-аферентите все още не е известно [Фиг. 1 (B2)], но са документирани нископрагови тактилни входове към спиноталамичните проекционни клетки,27 което дава достоверност на съобщенията за фини, контралатерални дефицити на откриване на докосване при пациенти след разрушаване на тези пътища след процедури на хордотомия.28
LTMRs в гладка кожа
Клетъчно-невритни комплекси на Меркел и сензорен купол. Меркел (1875) е първият, който дава хистологично описание на клъстери от епидермални клетки с големи лобулирани ядра, осъществяващи контакт с предполагаеми аферентни нервни влакна. Той предположи, че те подчиняват усещането за допир, като ги наричат Tastzellen (тактилни клетки). При хората клетъчните невритни комплекси на Меркел са обогатени в чувствителните на допир области на кожата, намират се в базалния слой на епидермиса на пръстите, устните и гениталиите. Те съществуват и в окосмена кожа с по-ниска плътност. Клетъчният невритен комплекс на Меркел се състои от клетка на Меркел, намираща се в тясна апозиция към увеличен нервен терминал от единичен миелинизиран A? влакно [фиг. 1 (C1)] (преглед в Halata и сътрудници).29 От епидермалната страна клетката на Меркел показва пръстовидни процеси, простиращи се между съседни кератиноцити [Фиг. 1 (C2)]. Клетките на Меркел са епидермални клетки, извлечени от кератиноцити.30,31 Терминът допирен купол е въведен за назоваване на голямата концентрация на клетъчни комплекси на Меркел в окосмената кожа на предната лапа на котката. Един сензорен купол може да има до 150 клетки на Меркел, инервирани от едно A?-влакно и при хората освен A?-влакна, A? и С-влакна също присъстваха редовно.32-34
Стимулирането на клетъчните невритни комплекси на Меркел води до бавно адаптиращи се отговори от тип I (SA I), които произлизат от прекъснати рецептивни полета с остри граници. Няма спонтанно изпускане. Тези комплекси реагират на дълбочината на вдлъбнатина в кожата и имат най-висока пространствена разделителна способност (0.5 mm) на кожните механорецептори. Те предават точно пространствено изображение на тактилни стимули и се предполага, че са отговорни за дискриминацията на формата и текстурата [Фиг. 2 (B1)]. Мишките, лишени от клетки на Меркел, не могат да открият текстурирани повърхности с краката си, докато го правят с помощта на мустаците си.35
Все още е въпрос на дебат дали клетката на Меркел, сензорният неврон или и двете са места на механотрансдукция. При плъхове фототоксичното разрушаване на клетките на Меркел премахва отговора на SA I.36 При мишки с генетично потиснати клетки на Меркел, отговорът на SA I, записан in ex vivo подготовка на кожата/нервите, напълно изчезва, демонстрирайки, че клетките на Меркел са необходими за правилното кодиране на Меркел рецепторни отговори.37 Въпреки това, механичната стимулация на изолирани клетки на Меркел в култура чрез двигателно задвижван натиск не генерира механично регулирани токове.38,39 Кератиноцитите могат да играят важна роля в нормалното функциониране на клетъчно-невритния комплекс на Меркел. Пръстоподобните процеси на клетките на Меркел могат да се движат с деформация на кожата и движение на клетките на епидермиса и това може да е първата стъпка от механичната трансдукция. Ясно е, че условията, необходими за изследване на механочувствителността на клетките на Меркел, все още не са установени.
Краища на Руфини. Краищата на Ruffini са тънки капсулирани сензорни окончания във формата на пура, свързани с A? нервни окончания. Краищата на Ruffini са малки съединителнотъканни цилиндри, разположени по протежение на дермални колагенови нишки, които се доставят от едно до три миелинизирани нервни влакна с диаметър 4 µm. До три цилиндъра с различна ориентация в дермата могат да се слеят, за да образуват един рецептор [фиг. 6 (C1)]. Структурно окончанията на Руфини са подобни на сухожилните органи на Голджи. Те са широко изразени в дермата и са идентифицирани като бавно адаптиращи се кожни механорецептори тип II (SA II). На фона на спонтанна нервна дейност, бавно адаптиращо се редовно отделяне се предизвиква от перпендикулярно механично стимулиране с ниска сила или по-ефективно чрез разтягане на кожата. Отговорът на SA II произлиза от големи рецептивни полета с неясни граници. Рецепторите на Ruffini допринасят за възприемането на посоката на движение на обекта чрез модела на разтягане на кожата [фиг. 3 (A2)].
При мишки отговорите на SA I и SA II могат да бъдат разделени електрофизиологично при препариране на нервна кожа ex vivo.40 Nandasena и сътрудници съобщават за имунолокализацията на аквапорин 1 (AQP1) в периодонталните Ruffini окончания на резците на плъх, което предполага, че AQP1 участва в поддържането на зъбния осмотичен баланс, необходим за механотрансдукцията.41 Пародонталните окончания на Ruffini също изразяват предполагаемия механочувствителен йонен канал ASIC3.42
Мейснеровите телца. Мейснеровите телца са локализирани в дермалните папили на голата кожа, главно в дланите на ръцете и стъпалата, но също и в устните, езика, лицето, зърната и гениталиите. Анатомично те се състоят от капсулиран нервен край, като капсулата е направена от сплескани поддържащи клетки, подредени като хоризонтални ламели, вградени в съединителната тъкан. Има ли едно нервно влакно А? аференти, свързани на корпускул [фиг. 1 (C4)]. Всяка физическа деформация на телцето предизвиква залп от потенциали на действие, които бързо спират, т.е. те са бързо адаптиращи се рецептори. Когато стимулът бъде премахнат, корпускулът възвръща своята форма и при това произвежда нов залп от потенциали на действие. Поради повърхностното си разположение в дермата, тези телца селективно реагират на движение на кожата, тактилно откриване на приплъзване и вибрации (20-40 Hz). Те са чувствителни към динамична кожа – например между кожата и обект, който се манипулира [фиг. 2 (A1)].
Пачинови телца. Телцата на Пацини са по-дълбоките механорецептори на кожата и са най-чувствителният капсулиран кожен механорецептор за движението на кожата. Тези големи яйцевидни телца (дължина 1 mm), направени от концентрични ламели от фиброзна съединителна тъкан и фибробласти, облицовани с плоски модифицирани Schwann клетки, се експресират в дълбоката дерма.43 В центъра на корпускула, в пълна с течност кухина, наречена вътрешна луковица , прекратява едно единствено A? аферентно немиелинизирано нервно окончание [фиг. 1 (C5)]. Те имат голямо рецептивно поле на повърхността на кожата с особено чувствителен център. Развитието и функцията на няколко бързо адаптиращи се типа механорецептори са нарушени при c-Maf мутантни мишки. По-специално, пациниевите телца са силно атрофирани.44
Телцата на Пацини показват много бърза адаптация в отговор на вдлъбнатината на кожата, бързо адаптиращия се II (RA II) нервен разряд, който е в състояние да следва висока честота на вибрационни стимули и позволява възприемане на далечни събития чрез предавани вибрации.45 Пациниан корпускул аферентите реагират на продължително вдлъбнатина с преходна активност в началото и отстраняването на стимула. Те се наричат още детектори за ускорение, защото могат да открият промени в силата на стимула и, ако скоростта на промяна в стимула се промени (както се случва при вибрациите), техният отговор става пропорционален на тази промяна. Телцата на Пачини усещат грубите промени в налягането и най-вече вибрациите (150-300 Hz), които могат да открият дори на сантиметри разстояние [фиг. 2 (A3)].
Тоническата реакция се наблюдава в декапсулирана пацинова телца.46 В допълнение, непокътнатите пацинови телца реагират с продължителна активност по време на постоянни стимули на вдлъбнатина, без да променят механичните прагове или честотата на реакция, когато GABA-медиираната сигнализация е блокирана между ламелатната глия и нервния край.47 По този начин, не-невронните компоненти на телцето на Пачин може да имат двойна роля във филтрирането на механичния стимул, както и в модулирането на свойствата на отговора на сензорния неврон.
Проекции на гръбначния мозък. Проекциите на A?-LTMRs в гръбначния мозък са разделени на два клона. Главният централен клон се издига в гръбначния мозък в ипсилатералните гръбни колони до нивото на шийката на матката [Фиг. 1 (B3)]. Вторичните разклонения завършват в дорзалния рог в ламината IV и пречат на предаването на болката, например. Това може да отслаби болката като част от контрола на портата [фиг. 1 (B4)].48
На нивата на шийката на матката аксоните на главния клон се разделят на два тракта: средната линия включва грацилния фаскул, предаващ информация от долната половина на тялото (крака и хобот), а външният тракт включва клиновидния пучок, предаващ информация от горната половина. на тялото (ръцете и багажника) [фиг. 1 (B5)].
Първичните тактилни аференти правят първия си синапс с неврони от втори порядък в медулата, където влакната от всеки тракт синапсират в едноименно ядро: синапсът на аксоните на gracile fasciculus в грацилното ядро и синапсът на клиновидните аксони в клиновидното ядро [Фиг. 1 (B6)]. Невроните, получаващи синапса, осигуряват вторичните аференти и незабавно пресичат средната линия, за да образуват тракт от контралатералната страна на мозъчния ствол – медиалния лемнискус – който се издига през мозъчния ствол до следващата релейна станция в средния мозък, по-специално в таламуса [Фиг. . 1 (B7)].
Молекулна спецификация на LTMR. Молекулните механизми, контролиращи ранната диверсификация на LTMR, наскоро бяха частично изяснени. Bourane и сътрудниците показват, че невронните популации, експресиращи рецептора на Ret тирозин киназа (Ret) и неговия ко-рецептор GFR?2 в E11 13 ембрионални мишки DRG, селективно коекспресират транскрипционния фактор Mafa.49,50 Тези автори демонстрират, че Mafa/ Ret/GFR?2 невроните, предназначени да се превърнат в три специфични типа LTRMs при раждането: невроните SA1, инервиращи комплекси от клетките на Меркел, бързо адаптиращите се неврони, инервиращи клетките на Мейснер, и бързо адаптиращите се аференти (RA I), образуващи ланцетни окончания около космените фоликули. Ginty и сътрудниците също така съобщават, че DRG невроните, експресиращи ранен Ret, бързо адаптират механорецептори от телцата на Майснер, пациниевите телца и ланцетните окончания около космените фоликули.51 Те инервират дискретни целеви зони в грацилните и клиновидни ядра, разкривайки модерен модел на механоса. невронни аксонални проекции в мозъчния ствол.
Изследване на механорецепторите на човешката кожа. Техниката на „микроневрографията“, описана от Hagbarth и Vallbo през 1968 г., е приложена за изследване на поведението на отделянето на отделни човешки механочувствителни краища, доставящи мускули, стави и кожа (вижте за преглед Macefield, 2005).52,53 По-голямата част от микроневрографията на човешката кожа проучвания характеризират физиологията на тактилните аференти в гладката кожа на ръката. Микроелектродните записи от медианните и улнарните нерви при хора разкриват усещане за допир, генерирано от четирите класа LTMR: аферентите на Meissner са особено чувствителни към леки удари по кожата, реагирайки на локални сили на срязване и начални или явни приплъзвания в рамките на рецептивното поле. Аферентите на Пачини са изключително чувствителни към бързи механични преходни процеси. Аферентите реагират енергично на издухване над рецептивното поле. Пацинова телца, разположена в една цифра, обикновено реагира на почукване по масата, поддържаща ръката. Аферентите на Меркел характерно имат висока динамична чувствителност към стимули за вдлъбнатина, приложени към дискретна област и често реагират с изключен разряд по време на освобождаване. Въпреки че аферентите на Ruffini наистина реагират на силите, приложени нормално към кожата, уникална характеристика на SA II аферентите е способността им да реагират и на странично разтягане на кожата. И накрая, космите в предмишницата имат големи яйцевидни или неправилни рецептивни полета, съставени от множество чувствителни петна, които съответстват на отделни косми (всяка аферентна доставка ~20 косъма).
Механична чувствителност на кератиноцитите
Всеки механичен стимул върху кожата трябва да се предава чрез кератиноцити, които образуват епидермиса. Тези повсеместни клетки могат да изпълняват сигнални функции в допълнение към техните поддържащи или защитни роли. Например, кератиноцитите секретират АТФ, важна сензорна сигнална молекула, в отговор на механични и осмотични стимули.54,55 Освобождаването на АТФ индуцира вътреклетъчно повишаване на калция чрез автокринна стимулация на пуринергичните рецептори.55 Освен това има доказателства, че хипотоничността активира Rho -киназен сигнален път и последващото образуване на F-актин стрес влакно, което предполага, че механичната деформация на кератиноцитите може механично да попречи на съседните клетки като клетките на Меркел за безвредно докосване и свободни краища на C-фибри за вредно докосване [Фиг. 1 (C6)].56,57
Вредно докосване
Механорецепторите с висок праг (HTMR) са епидермални C- и A? свободни нервни окончания. Те не са свързани със специализирани структури и се наблюдават както в окосмена кожа [фиг. 1 (A9)] и гола кожа [фиг. 1(C7)]. Въпреки това, терминът на свободните нервни окончания трябва да се разглежда разумно, тъй като нервните окончания винаги са в тясна връзка с кератиноцитите или клетките на Langherans или меланоцитите. Ултраструктурният анализ на нервните окончания разкрива наличието на груб ендоплазмен ретикулум, изобилни митохондрии и плътно-ядрени везикули. Съседните мембрани на епидермалните клетки са удебелени и наподобяват постсинаптична мембрана в нервните тъкани. Имайте предвид, че взаимодействията между нервните окончания и епидермалните клетки могат да бъдат двупосочни, тъй като епидермалните клетки могат да освобождават медиатори като АТФ, интерлевкин (IL6, IL10) и брадикинин и обратно пептидергичните нервни окончания могат да освобождават пептиди като CGRP или вещество Р, действащи върху епидермалните клетки. HTMR включват механо-ноцицептори, възбудени само от вредни механични стимули и полимодални ноцицептори, които също реагират на вредна топлина и екзогенен химикал [Фиг. 2 (B2)].58
HTMR аферентните влакна завършват на проекционните неврони в дорзалния рог на гръбначния мозък. Ap-HTMRs контактуват с неврони от втори ред предимно в ламина I и V, докато C-HTMRs завършват в ламина II [Фиг. 1 (B8)]. Ноцицептивните неврони от втори ред се проектират към контролната страна на гръбначния мозък и се издигат в бялото вещество, образувайки предностранната система. Тези неврони завършват главно в таламуса [фиг. 1 (B9 и B10)].
Механо-токове в соматосензорните неврони
Механизмите на бавна или бърза адаптация на механорецепторите все още не са изяснени. Не е ясно до каква степен адаптацията на механорецепторите се осигурява от клетъчната среда на сетивните нервни окончания, присъщите свойства на механично затворените канали и свойствата на аксоналните волтаж-зависими йонни канали в сензорните неврони (фиг. 2). Въпреки това, скорошният напредък в характеризирането на механично затворени токове показа, че различни класове механочувствителни канали съществуват в DRG невроните и могат да обяснят някои аспекти на адаптацията на механорецепторите.
Записването in vitro при гризачи показва, че сомата на DRG невроните е присъщо механочувствителна и експресира катионни механо-зависими токове.59-64 Гадолиниевото и рутениевото червено напълно блокират механочувствителните токове, докато външни калций и магнезий, при физиологични концентрации, както и амилорид и бензамил, причиняват частичен блок.60,62,63 FM1-43 действа като траен блокер, а инжектирането на FM1-43 в задната лапа на мишки намалява чувствителността към болка в теста на Randall-Selitto и повишава оценения праг на изтегляне на лапата с косми на фон Фрей.65
В отговор на продължителна механична стимулация, механочувствителните токове намаляват чрез затваряне. Въз основа на времевите константи на затихването на тока са разграничени четири различни типа механочувствителни токове: бързо адаптиращи се токове (~3 ms), междинно адаптиращи се токове (~6 ms), бавно адаптиращи се токове (~15 ms ) и ултра-бавно адаптиращи се токове (~30 ms).200 Всички тези токове присъстват с променлива честота в DRG невроните на плъхове, инервиращи голата кожа на задната лапа.300
Механичната чувствителност на механочувствителните токове може да бъде определена чрез прилагане на серия от нарастващи механични стимули, позволяващи сравнително подробен анализ стимул-ток.66 Връзката между стимул-ток обикновено е сигмоидална, а максималната амплитуда на тока се определя от броя на канали, които са едновременно отворени.64,67 Интересно е, че бързо адаптиращият се механочувствителен ток показва нисък механичен праг и средна точка на полуактивиране в сравнение с ултра бавно адаптиращия се механочувствителен ток.63,65
Сензорните неврони с не-ноцицептивни фенотипове за предпочитане експресират бързо адаптиращи се механочувствителни токове с по-нисък механичен праг.60,61,63,64,68 Обратно, бавно и ултра-бавно адаптиращи се механочувствителни токове понякога се съобщават в предполагаеми неноцицептивни клетки.64,68 Това предизвика предположение, че тези токове могат да допринесат за различните механични прагове, наблюдавани в LTMR и HTMR in vivo. Въпреки че тези in vitro експерименти трябва да се предприемат с повишено внимание, подкрепата за присъствието в сомата на DRG невроните на механотрансдюсери с нисък и висок праг беше предоставена и чрез радиално разтягане на стимулиране на култивирани миши сензорни неврони.69 Тази парадигма разкрива две основни популации от чувствителни към разтягане неврони, една, която реагира на ниска амплитуда на стимула, и друга, която селективно реагира на висока амплитуда на стимула.
Тези резултати имат важни, но спекулативни, механистични последици: механичният праг на сензорните неврони може да има малко общо с клетъчната организация на механорецептора, но може да се крие в свойствата на механично затворените йонни канали.
Механизмите, които са в основата на десенсибилизацията на механочувствителните катионни токове в DRG невроните на плъхове, бяха наскоро разкрити.64,67 Това е резултат от два едновременни механизма, които влияят на свойствата на канала: адаптация и инактивация. Адаптацията е съобщена за първи път в проучвания на слухови космени клетки. Може да се опише оперативно като прост транслация на кривата на активиране на канала на трансдюсера по оста на механичния стимул.70-72 Адаптирането позволява на сензорните рецептори да поддържат чувствителността си към нови стимули в присъствието на съществуващ стимул. Въпреки това, значителна част от механочувствителните токове в DRG невроните не могат да бъдат реактивирани след кондициониране на механична стимулация, което показва инактивиране на някои трансдюсерни канали.64,67 Следователно, както инактивацията, така и адаптацията действат в тандем за регулиране на механочувствителните токове. Тези два механизма са общи за всички механочувствителни токове, идентифицирани в DRG невроните на плъхове, което предполага, че свързаните физикохимични елементи определят кинетиката на тези канали.64
В заключение, определянето на свойствата на ендогенните механочувствителни токове in vitro е от решаващо значение в стремежа да се идентифицират механизмите на трансдукция на молекулярно ниво. Променливостта, наблюдавана в механичния праг и адаптивната кинетика на различните механично-зависими токове в DRG невроните предполагат, че присъщите свойства на йонните канали могат да обяснят, поне отчасти, механичния праг и адаптационната кинетика на механорецепторите, описани през десетилетията от 1960 г. 80, използвайки препарати ex vivo.
Предполагаеми механочувствителни протеини
Механочувствителните йонни потоци в соматосензорните неврони са добре характеризирани, за разлика от тях, малко се знае за идентичността на молекулите, които медиират механотрансдукцията при бозайници. Генетичните скрининги в Drosophila и C. elegans идентифицират кандидат-механотрансдукционни молекули, включително семействата TRP и дегенеринови/епителни Na+ канали (Deg/ENaC).73 Последните опити за изясняване на молекулярната основа на механотрансдукцията при бозайници до голяма степен се фокусират върху тези кандидат-хомолози . Освен това много от тези кандидати присъстват в кожните механорецептори и соматосензорните неврони (фиг. 2).
Йонни канали, чувствителни към киселина
ASICs принадлежат към протон-зависима подгрупа от семейството на дегенеринови епителни Na+ канали.74 Три члена от семейството ASIC (ASIC1, ASIC2 и ASIC3) се експресират в механорецептори и ноцицептори. Ролята на ASIC каналите е изследвана в поведенчески проучвания, използващи мишки с целенасочена делеция на гените на ASIC канала. Изтриването на ASIC1 не променя функцията на кожните механорецептори, но повишава механичната чувствителност на аферентите, инервиращи червата.75 ASIC2 нокаут мишките показват намалена чувствителност на бързо адаптиращите се кожни LTMR.76 Въпреки това, последващи проучвания съобщават за липса на ефекти от нокаутирането на ASIC2 върху както висцерална механо-ноцицепция, така и кожна механосензация.77 Разрушаването на ASIC3 намалява механочувствителността на висцералните аференти и намалява отговорите на кожните HTMR на вредни стимули.76
Преходният рецепторен канал
TRP суперсемейството е подразделено на шест подсемейства при бозайници.78 Почти всички TRP подсемейства имат членове, свързани с механочувствителност в различни клетъчни системи.79 При сензорните неврони на бозайниците обаче TRP каналите са най-известни с усещането на термична информация и медиирането на неврогенно възпаление, и само два TRP канала, TRPV4 и TRPA1, са замесени в реакцията на докосване. Нарушаването на експресията на TRPV4 при мишки има само скромни ефекти върху острите механосензорни прагове, но силно намалява чувствителността към вредни механични стимули.80,81 TRPV4 е решаващ определящ фактор при оформянето на отговора на ноцицептивните неврони към осмотичен стрес и към механична хипералгезия.82,83 по време на инфлация,1mmmm 1 TRPA1 изглежда има роля в механичната хипералгезия. TRPA84,85-дефицитни мишки проявяват свръхчувствителност към болка. TRPAXNUMX допринася за трансдукцията на механични, студени и химични стимули в ноцицепторните сензорни неврони, но изглежда, че това не е от съществено значение за трансдукцията на космените клетки.XNUMX
Няма ясни доказателства, показващи, че TRP каналите и ASIC каналите, експресирани при бозайници, са механично затворени. Нито един от тези канали, изразени хетерологично, не рекапитулира електрическия подпис на механочувствителните токове, наблюдавани в тяхната естествена среда. Това не изключва възможността ASIC и TRPs каналите да са механотрансдюсери, като се има предвид несигурността дали канал за механотрансдукция може да функционира извън своя клетъчен контекст (вижте раздел за SLP3).
Пиезо протеини
Пиезо протеините наскоро бяха идентифицирани като обещаващи кандидати за механосензиращи протеини от Кост и сътрудници.86,87 Гръбначните имат два члена на Piezo, Piezo 1 и Piezo 2, по-рано известни като FAM38A и FAM38B, съответно, които са добре запазени в многоклетъчния еук . Piezo 2 е в изобилие в DRGs, докато Piezo 1 е едва откриваем. Пиезоиндуцираните механочувствителни токове се предотвратяват инхибирани от гадолиний, рутениев червен и GsMTx4 (токсин от тарантулата Grammostola spatulata).88 Експресията на Piezo 1 или Piezo 2 в хетероложни системи произвежда механочувствителни токове, като кинетиката на Piezoactiv2 е бърза кинетика на тока отколкото Piezo 1. Подобно на ендогенните механочувствителни токове, пиезозависимите токове имат обратен потенциал около 0 mV и не са катионно селективни, като Na+, K+, Ca2+ и Mg2+ всички проникват в основния канал. По същия начин, пиезозависимите токове се регулират от мембранния потенциал, с подчертано забавяне на кинетиката на тока при деполяризирани потенциали.86
Пиезо протеините несъмнено са механочувствителни протеини и споделят много свойства на бързо адаптиращи се механочувствителни токове в сензорните неврони. Третирането на култивирани DRG неврони с Piezo 2 къса интерферираща РНК намалява дела на невроните с бързо адаптиращ се ток и намалява процента на механочувствителните неврони.86 Трансмембранните домени са разположени в пиезо протеините, но не са открити очевидни мотиви, съдържащи пори или сигнатури на йонни канали. идентифициран. Въпреки това, миши Piezo 1 протеин, пречистен и възстановен в асиметрични липидни бислоеве и липозоми, образува йонни канали, чувствителни към рутениево червено.87 Съществена стъпка в валидирането на механотрансдукцията чрез пиезо каналите е използването на in vivo подходи за определяне на функционалното значение при сигнализирането на докосване. Информацията беше дадена в Drosophila, където изтриването на единичния член на Piezo намалява механичния отговор на вредни стимули, без да засяга нормалното докосване.89 Въпреки че тяхната структура предстои да бъде определена, това ново семейство от механочувствителни протеини е обещаващ предмет за бъдещи изследвания, отвъд границата на усещане за докосване. Например, скорошно проучване върху пациенти с анемия (наследствена ксероцитоза) показва ролята на Piezo 1 в поддържането на хомеостазата на обема на еритроцитите.90
Подобен на трансмембранен канал (TMC)
Едно скорошно проучване показва, че два протеина, TMC1 и TMC2, са необходими за механотрансдукция на космените клетки.91 Наследствена глухота, дължаща се на TMC1 генна мутация, е докладвана при хора и мишки.92,93 Наличието на тези канали все още не е показано в соматосензорната система , но изглежда е добър повод за разследване.
Стоматин-подобен протеин 3 (SLP3)
В допълнение към трансдукционните канали е показано, че някои спомагателни протеини, свързани с канала, играят роля в чувствителността на докосване. SLP3 се експресира в DRG неврони на бозайници. Изследвания, използващи мутантни мишки без SLP3, показват промяна в механочувствителност и механочувствителни токове.94,95 Точната функция на SLP3 остава неизвестна. Може да е линкер между механочувствителния канал и подлежащите микротубули, както е предложено за неговия хомолог на C. elegans MEC2.96 Наскоро GR. Лабораторията на Lewin предполага, че тетерът се синтезира от DRG сензорните неврони и свързва механочувствителния йонен канал с извънклетъчния матрикс.97 Разрушаването на връзката премахва RA-механочувствителния ток, което предполага, че някои йонни канали са механочувствителни само когато са свързани. RA-механочувствителните течения също се инхибират от ламинин-332, матричен протеин, произведен от кератиноцити, засилвайки хипотезата за модулация на механочувствителния ток от извънклетъчни протеини.98
Подфамилия K+ канали
Успоредно с катионните деполяризиращи механочувствителни токове се изследва наличието на реполяризиращи механочувствителни K+ токове. K+ каналите в механочувствителните клетки могат да влязат в текущия баланс и да допринесат за определяне на механичния праг и времевия ход на адаптация на механорецепторите.
Членовете на KCNK принадлежат към семейството на K+ канали с две пори (K2P).99,100 K2P показва забележителен диапазон на регулиране от клетъчни, физически и фармакологични агенти, включително промени в pH, топлина, разтягане и деформация на мембраната. Тези K2P са активни при покой на мембранния потенциал. Няколко KCNK субединици се експресират в соматосензорни неврони.101 KCNK2 (TREK-1), KCNK4 (TRAAK) и TREK-2 канали са сред малкото канали, за които е показано директно механично стробиране чрез разтягане на мембраната.102,103
Мишките с нарушен KCNK2 ген показват повишена чувствителност към топлина и леки механични стимули, но нормален праг на изтегляне към вредно механично налягане, приложено към задната лапа с помощта на теста на Randall-Selitto.104 KCNK2-дефицитните мишки също показват повишена термична и механична хипералгезия при възпалителни процеси. условия. KCNK4 нокаут мишки са свръхчувствителни към лека механична стимулация и тази свръхчувствителност се увеличава чрез допълнително инактивиране на KCNK2.105 Повишената механочувствителност на тези нокаут мишки може да означава, че разтягането нормално активира както деполяризиращи, така и реполяризиращи механочувствителни токове по подобен на некоординиран начин деполяризиращи и реполяризиращи напрежение-зависими токове.
KCNK18 (TRESK) е основен принос за фоновата K+ проводимост, която регулира потенциала на мембраната в покой на соматосензорните неврони.106 Въпреки че не е известно дали KCNK18 е пряко чувствителен към механична стимулация, той може да играе роля в медиирането на реакциите на леко докосване, както и болезнени механични стимули. Предполага се, че KCNK18 и в по-малка степен KCNK3 са молекулярната мишена на хидрокси-?-sanshool, съединение, намерено в зърната пипер на Шезуан, което активира рецепторите за докосване и предизвиква усещане за изтръпване при хората.107,108
Зависимият от напрежението K+ канал KCNQ4 (Kv7.4) е от решаващо значение за определяне на предпочитанията за скорост и честота на субпопулация от бързо адаптиращи се механорецептори както при мишки, така и при хора. Мутацията на KCNQ4 първоначално е била свързана с форма на наследствена глухота. Интересно е, че едно скорошно проучване локализира KCNQ4 в периферните нервни окончания на кожния бързо адаптиращ се космен фоликул и телцето на Мейснер. Съответно, загубата на KCNQ4 функция води до селективно повишаване на чувствителността на механорецепторите към нискочестотни вибрации. По-специално, хората с късна загуба на слуха, дължаща се на доминантни мутации на гена KCNQ4, показват подобрена ефективност при откриване на вибрации с малка амплитуда, нискочестотни.109
Прозрение на д-р Алекс Химензе
Докосването се счита за едно от най-сложните сетива в човешкото тяло, особено защото няма специфичен орган, който да отговаря за него. Вместо това, усещането за допир се осъществява чрез сензорни рецептори, известни като механорецептори, които се намират в кожата и реагират на механичен натиск или изкривяване. Има четири основни типа механорецептори в голата или безкосмена кожа на бозайниците: ламеларни корпускули, тактилни телца, нервни окончания на Меркел и луковични телца. Механорецепторите функционират, за да позволят откриването на докосване, за да следят позицията на мускулите, костите и ставите, известни като проприоцепция, и дори за откриване на звуци и движение на тялото. Разбирането на механизмите на структурата и функцията на тези механорецептори е основен елемент в използването на лечения и терапии за управление на болката.
Заключение
Докосването е сложно усещане, защото представя различни тактилни качества, а именно вибрация, форма, текстура, удоволствие и болка, с различни дискриминационни изпълнения. Досега съответствието между сензорния орган и психофизичния усет беше корелативно и специфичните за класа молекулярни маркери тепърва се появяват. Разработването на тестове за гризачи, съответстващи на разнообразието от поведение на докосване, сега е необходимо, за да се улесни бъдещата идентификация на геномиката. Използването на мишки, при които липсват специфични подгрупи от сензорни аферентни типове, значително ще улесни идентифицирането на механорецепторите и сензорните аферентни влакна, свързани с определен начин на докосване. Интересно е, че неотдавнашна статия отваря важния въпрос за генетичната основа на механосензорните черти при човека и предполага, че единична генна мутация може да повлияе негативно на чувствителността на докосване.110 Това подчертава, че патофизиологията на дефицита на допир при човека е до голяма степен неизвестна и със сигурност би напредък чрез идентифициране точно на подмножеството от сензорни неврони, свързани с модалност на докосване или дефицит на докосване.
В замяна на това е постигнат напредък при дефинирането на биофизичните свойства на механизираните токове.64 Развитието на нови техники през последните години, позволяващи наблюдение на промените в мембранното напрежение при записване на механо-регулирания ток, се оказа ценен експериментален метод за описване механочувствителни токове с бърза, междинна и бавна адаптация (прегледани в Delmas и сътрудници).66,111 Бъдещето ще бъде да се определи ролята на текущите свойства в механизмите на адаптация на функционално различни механорецептори и приноса на механочувствителните K+ токове към възбудимостта на LTMR и HTMR.
Молекулната природа на механизираните токове при бозайници също е бъдеща обещаваща изследователска тема. Бъдещите изследвания ще напредват в две перспективи, първо за определяне на ролята на спомагателната молекула, която свързва каналите към цитоскелета и ще бъде необходима за придаване или регулиране на механочувствителността на йонните канали от семействата TRP и ASIC/EnaC. Второ, да се изследва голямата и обещаваща област на приноса на пиезо каналите, като се отговори на ключови въпроси, свързани с механизмите за проникване и стробиране, подмножеството от сензорни неврони и докосни модалности, включващи Piezo и ролята на Piezo в неневронни клетки, свързани с механосенсетивност.
Усещането за допир, в сравнение с това на зрението, вкуса, звука и обонянието, които използват специфични органи за обработка на тези усещания, може да се появи в цялото тяло чрез малки рецептори, известни като механорецептори. Различни видове механорецептори могат да бъдат намерени в различни слоеве на кожата, където могат да открият широк спектър от механична стимулация. Статията по-горе описва конкретни акценти, които демонстрират напредъка на структурните и функционални механизми на механорецепторите, свързани с усещането за допир. Информация от Националния център за биотехнологична информация (NCBI). Обхватът на нашата информация е ограничен до хиропрактика, както и до наранявания и състояния на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителни теми: Болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати дни по време на работа в световен мащаб. Всъщност, болката в гърба е приписана като втората най-често срещана причина за посещенията в лекарски кабинети, превишаващи броя им само чрез инфекции на горните дихателни пътища. Приблизително 80 процента от населението ще преживее някакъв вид болка в гърба поне веднъж през целия си живот. Гърбът е сложна структура, съставена от кости, стави, връзки и мускули, сред другите меки тъкани. Поради това, наранявания и / или влошени условия, като например херния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
1. Moriwaki K, Yuge O. Топографски характеристики на кожни тактилни хипоестетични и хиперестетични аномалии при хронична болка.Болка. 1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Крос Реф]
2. Shim B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Механична и топлинна сенсибилизация на кожни ноцицептори при плъхове с експериментална периферна невропатия.Невронаука. 2005;132:193 201. doi: 10.1016/j.neuroscience.2004.12.036. [PubMed]�[Крос Реф]
3. Kleggetveit IP, J rum E. Дисфункция на големи и малки влакна при наранявания на периферните нерви със или без спонтанна болка. J Pain. 2010;11:1305 10. doi: 10.1016/j.jpain.2010.03.004. [PubMed]�[Крос Реф]
4. Noback CR. Морфология и филогения на косата. Ann NY Acad Sci. 1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed]�[Крос Реф]
5. Straile НИЕ. Атипични предпазни космени фоликули в кожата на заека. Природа. 1958;181:1604�5. doi: 10.1038/1811604a0.�[PubMed]�[Крос Реф]
6. Straile НИЕ. Морфологията на тилотриховите фоликули в кожата на заека. Am J Anat. 1961;109:1 13. doi: 10.1002/aja.1001090102. [PubMed]�[Крос Реф]
7. Милард CL, Улф CJ. Сензорна инервация на космите на задния крайник на плъх: светлинен микроскопски анализ. J Comp Neurol. 1988;277: 183 94. doi: 10.1002/cne.902770203. [PubMed]�[Крос Реф]
8. Hamann W. Кожни механорецептори на бозайници. Prog Biophys Mol Biol. 1995;64:81�104. doi: 10.1016/0079-6107(95)00011-9.�[Review]�[PubMed]�[Крос Реф]
9. Brown AG, Iggo A. Количествено изследване на кожни рецептори и аферентни влакна при котка и заек.J Physiol. 1967;193:707 33. [PMC безплатна статия]�[PubMed]
10. Burgess PR, Petit D, Warren RM. Типове рецептори в котешка окосмена кожа, снабдени от миелинизирани влакна. J Неврофизиол. 1968;31:833 48. [PubMed]
11. Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. Sox2-позитивните дермални папила клетки определят вида на космения фоликул в епидермиса на бозайници.Развитие. 2009;136:2815 23. doi: 10.1242/dev.038620. [PMC безплатна статия]�[PubMed]�[Крос Реф]
12. Хюсеин М.А. Цялостният модел на подреждане на космения фоликул при плъхове и мишки. J Anat. 1971;109:307 16. [PMC безплатна статия]�[PubMed]
13. Vielkind U, Hardy MH. Промяна на моделите на молекулите на клетъчната адхезия по време на развитието на космения фоликул на мишките кожички. 2. Морфогенеза на фоликула при мутанти на косата, таби и пух. Acta Anat (Базел) 1996;157:183 94. doi: 10.1159/000147880. [PubMed]�[Крос Реф]
14. Hardy MH, Vielkind U. Промяна на моделите на молекулите на клетъчна адхезия по време на развитието на космения фоликул на мишките кожички. 1. Фоликулна морфогенеза при мишки от див тип. Acta Anat (Базел) 1996;157:169 82. doi: 10.1159/000147879. [PubMed]�[Крос Реф]
15. Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S и др. Функционалната организация на кожните нископрагови механосензорни неврони. Клетка. 2011;147:1615 27. doi: 10.1016/j.cell.2011.11.027.[PMC безплатна статия]�[PubMed]�[Крос Реф]
16. Brown AG, Iggo A. Количествено изследване на кожни рецептори и аферентни влакна при котка и заек.J Physiol. 1967;193:707 33. [PMC безплатна статия]�[PubMed]
17. Burgess PR, Petit D, Warren RM. Типове рецептори в котешка окосмена кожа, снабдени от миелинизирани влакна. J Неврофизиол. 1968;31:833 48. [PubMed]
18. Vallbo A, Olausson H, Wessberg J, Norrsell U. Система от немиелинизирани аференти за безвредна механорецепция в човешката кожа.Brain Res. 1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed]�[Крос Реф]
19. Vallbo AB, Olausson H, Wessberg J. Немиелинизираните аференти представляват втора система, кодираща тактилни стимули на човешката окосмена кожа.J Неврофизиол. 1999;81:2753 63. [PubMed]
21. McGlone F, Vallbo AB, Olausson H, Loken L, Wessberg J. Дискриминативно докосване и емоционално докосване.Може ли J Exp Psychol. 2007;61:173 83. doi: 10.1037/cjep2007019. [PubMed]�[Крос Реф]
22. Wessberg J, Olausson H, Fernstrm KW, Vallbo AB. Рецептивни полеви свойства на немиелинизирани тактилни аференти в човешката кожа. J Неврофизиол. 2003;89:1567 75. doi: 10.1152/jn.00256.2002. [PubMed][Крос Реф]
23. Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Молекулярно-генетична визуализация на рядка подгрупа от немиелинизирани сензорни неврони, които могат да открият нежно докосване.Nat Neurosci.2007;10:946 8. doi: 10.1038/nn1937. [PubMed]�[Крос Реф]
24. Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G и др. Немиелинизирани тактилни аференти сигнализират за докосване и проектират към островната кора. Nat Neurosci.2002;5:900 4. doi: 10.1038/nn896.[PubMed]�[Крос Реф]
25. Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. Неврофизиологията на немиелинизираните тактилни аференти.Neurosci Biobehav Rev. 2010;34: 185. doi: 91/j.neubiorev.10.1016. [Преглед][PubMed]�[Крос Реф]
26. Kr mer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M и др. Активиране на кортикалната мрежа за болка чрез мека тактилна стимулация след инжектиране на суматриптан.Болка. 2007;133:72 8. doi: 10.1016/j.pain.2007.03.001. [PubMed]�[Крос Реф]
27. Applebaum AE, Beall JE, Foreman RD, Willis WD. Организация и рецептивни полета на невроните на спиноталамичния тракт на приматите. J Неврофизиол. 1975;38:572 86. [PubMed]
28. Бяло JC, Sweet WH. Ефективност на хордотомията при фантомна болка след ампутация. AMA Arch Neurol Psychiatry. 1952;67:315 22. [PubMed]
29. Халата З, Грим М, Бауман К.И. Фридрих Зигмунд Меркел и неговата „клетка на Меркел“, морфология, развитие и физиология: преглед и нови резултати.Anat Rec A Discov Mol Cell Evol Biol. 2003;271: 225. doi: 39/ar.a.10.1002. [PubMed]�[Крос Реф]
30. Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Клетките на Меркел от бозайници произлизат от епидермалната линия. Dev Biol. 2009;336:76. doi: 83/j.ydbio.10.1016.[PMC безплатна статия]�[PubMed]�[Крос Реф]
31. Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N и др. Епидермалните прогенитори пораждат клетките на Меркел по време на ембрионалното развитие и хомеостазата при възрастни.J Cell Biol. 2009;187: 91. doi: 100/jcb.10.1083. [PMC безплатна статия]�[PubMed]�[Крос Реф]
32. Ebara S, Kumamoto K, Baumann KI, Halata Z. Триизмерните анализи на докосни куполи в окосмената кожа на котешка лапа разкриват морфологични субстрати за сложна сензорна обработка. Neurosci Res. 2008;61: 159. doi: 71/j.neures.10.1016. [PubMed]�[Крос Реф]
33. Guinard D, Usson Y, Guillermet C, Saxod R. Merkel Комплекси от човешка цифрова кожа: триизмерно изображение с конфокална лазерна микроскопия и двойна имунофлуоресценция. J Comp Neurol. 1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Крос Реф]
34. Reinisch CM, Tschachler E. Сензорният купол в човешката кожа се доставя от различни видове нервни влакна. Ан Нейрол.2005;58: 88. doi: 95/ana.10.1002. [PubMed]�[Крос Реф]
35. Maricich SM, Morrison KM, Mathes EL, Brewer BM. Гризачите разчитат на клетките на Меркел за задачи за дискриминация на текстурата. J Neurosci. 2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[PMC безплатна статия]�[PubMed]�[Крос Реф]
36. Ikeda I, Yamashita Y, Ono T, Ogawa H. Селективното фототоксично унищожаване на клетките на Меркел от плъх премахва реакциите на бавно адаптиращите се механорецепторни единици тип I.J Physiol. 1994;479:247 56. [PMC безплатна статия][PubMed]
37. Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA и др. Клетките на Меркел са от съществено значение за реакциите на леко докосване. Наука. 2009;324:1580 2. doi: 10.1126/наука.1172890.[PMC безплатна статия]�[PubMed]�[Крос Реф]
38. Diamond J, Holmes M, медицинска сестра CA. Участват ли реципрочни синапси на клетъчно-невритни клетки на Меркел в инициирането на тактилни реакции в кожата на саламандър?J Physiol. 1986;376:101 20. [PMC безплатна статия][PubMed]
39. Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Токове, зависими от напрежението в изолирани единични клетки на Меркел на плъхове. J Physiol. 1992;450:143 62. [PMC безплатна статия]�[PubMed]
40. Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. Редовността на продължително изстрелване разкрива две популации от бавно адаптиращи се рецептори за докосване в космата кожа на мишката.J Неврофизиол. 2010;103:3378 88. doi: 10.1152/jn.00810.2009. [PMC безплатна статия]�[PubMed]�[Крос Реф]
41. Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Имунолокализация на аквапорин-1 в механорецептивните Ruffini окончания в пародонталния лигамент. Brain Res. 2007;1157:32. doi: 40/j.brainres.10.1016. [PubMed]�[Крос Реф]
42. Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K и др. Откриване на киселинно-чувствителен йонен канал 3 (ASIC3) в периодонталните Ruffini окончания на миши резци. Neurosci Lett. 2011;488:173 7. doi: 10.1016/j.neulet.2010.11.023. [PubMed]�[Крос Реф]
43. КО на Джонсън. Ролите и функциите на кожните механорецепториCurr Opin Neurobiol. 2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed]�[Крос Реф]
44. Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A и др. Транскрипционният фактор c-Maf контролира развитието и функцията на рецепторите за докосване. Наука. 2012;335:1373 6. doi: 10.1126/science.1214314. [PubMed]�[Крос Реф]
45. Менделсън М, Лоуенщайн WR. Механизми на рецепторна адаптация. Наука. 1964;144:554 5. doi: 10.1126/science.144.3618.554. [PubMed]�[Крос Реф]
46. Loewenstein WR, Mendelson M. Компоненти на рецепторната адаптация в пацинова телца. J Physiol. 1965;177:377 97. [PMC безплатна статия]�[PubMed]
47. Pawson L, Prestia LT, Mahoney GK, G l B, Cox PJ, Pack AK. ГАМКергичното/глутаматергично-глиално/невронно взаимодействие допринася за бързата адаптация в пациниановите телца. J Neurosci. 2009;29:2695�705. doi: 10.1523/JNEUROSCI.5974-08.2009.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
48. Basbaum AI, Jessell TM. Възприятието за болка. В: Kandel ER, Schwartz JH, Jessell TM, eds. Принципи на невронната наука. Четвърто издание. The McGraw-Hill compagies, 2000: 472-490.
49. Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A и др. Подтиповете на механорецепторите с нисък праг селективно експресират MafA и се определят чрез Ret сигнализиране. Неврон. 2009;64: 857. doi: 70/j.neuron.10.1016. [PubMed]�[Крос Реф]
50. Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. Роля за сигнализирането на транскрипционния фактор на Runx в диверсификацията на сензорните неврони на дорзалния корен на ганглия. Неврон. 2006;49: 379. doi: 93/j.neuron.10.1016. [PubMed]�[Крос Реф]
51. Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Молекулярна идентификация на бързо адаптиращи се механорецептори и зависимостта им от развитието на ret сигнализация. Неврон. 2009;64: 841. doi: 56/j.neuron.10.1016. [PMC безплатна статия]�[PubMed]�[Крос Реф]
52. Vallbo AB, Hagbarth KE. Активност от кожни механорецептори, регистрирана перкутанно при будни хора. Exp Neurol. 1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed]�[Крос Реф]
53. Macefield VG. Физиологични характеристики на нископраговите механорецептори в ставите, мускулите и кожата при хора. Clin Exp Pharmacol Physiol. 2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed]�[Крос Реф]
54. Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Ca2+ вълните в кератиноцитите се предават на сензорните неврони: участието на извънклетъчния ATP и P2Y2 рецепторното активиране. Biochem J. 2004;380:329 38. doi: 10.1042/BJ20031089. [PMC безплатна статия]�[PubMed][Крос Реф]
55. Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. Сигнализацията на ATP е от решаващо значение за отговора на човешките кератиноцити на механична стимулация чрез хипоосмотичен шок. Exp Dermatol. 2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed]�[Крос Реф]
56. Amano M, Fukata Y, Kaibuchi K. Регулация и функции на Rho-асоциираната киназа. Exp Cell Res. 2000;261:44. doi: 51/excr.10.1006. [Преглед] [PubMed]�[Крос Реф]
57. Koyama T, Oike M, Ito Y. Участие на Rho-киназа и тирозин киназа в индуцираното от хипотоничен стрес освобождаване на АТФ в ендотелните клетки на говедата аорта.J Physiol. 2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
58. Perl ER. Кожни полимодални рецептори: характеристики и пластичност. Prog Brain Res. 1996;113:21�37. doi: 10.1016/S0079-6123(08)61079-1.�[Review]�[PubMed]�[Крос Реф]
59. McCarter GC, Reichling DB, Levine JD. Механична трансдукция от ганглийни неврони на гръбначния корен на плъх in vitro. Neurosci Lett. 1999;273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[PubMed]�[Крос Реф]
60. Drew LJ, Wood JN, Cesare P. Различни механочувствителни свойства на чувствителни към капсаицин и нечувствителни сензорни неврони.J Neurosci. 2002;22:RC228. [PubMed]
61. Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P и др. Чувстващите киселини йонни канали ASIC2 и ASIC3 не допринасят за механично активирани токове в сензорните неврони на бозайници.J Physiol. 2004;556:691 710. doi: 10.1113/jphysiol.2003.058693. [PMC безплатна статия]�[PubMed]�[Крос Реф]
62. McCarter GC, Levine JD. Йонна основа на механотрансдукционния ток в невроните на ганглийния дорзален корен на възрастни плъхове.Mol Pain. 2006;2:28. doi: 10.1186/1744-8069-2-28.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
63. Coste B, Crest M, Delmas P. Фармакологична дисекция и разпределение на NaN/Nav1.9, Т-тип Ca2+ токове и механично активирани катионни токове в различни популации на DRG неврони. J Gen Physiol. 2007;129:57. doi: 77/jgp.10.1085. [PMC безплатна статия]�[PubMed]�[Крос Реф]
64. Hao J, Delmas P. Множество механизми за десенсибилизация на каналите на механотрансдюсерите оформят изстрелването на механосензорните неврони.J Neurosci. 2010;30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[PubMed]�[Крос Реф]
65. Дрю LJ, Wood JN. FM1-43 е перманентен блокер на механочувствителните йонни канали в сензорните неврони и инхибира поведенческите реакции към механични стимули.Mol Pain. 2007;3:1. doi: 10.1186/1744-8069-3-1.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
66. Hao J, Delmas P. Записване на механочувствителни токове с помощта на пиезоелектрически задвижван механостимулатор. Nat Protoc. 2011;6: 979. doi: 90/nprot.10.1038. [PubMed]�[Крос Реф]
67. Rugiero F, Drew LJ, Wood JN. Кинетични свойства на механично активираните токове в гръбначните сензорни неврони. J Physiol. 2010;588:301 14. doi: 10.1113/jphysiol.2009.182360. [PMC безплатна статия]�[PubMed][Крос Реф]
68. Ху Дж, Люин GR. Механочувствителни токове в невритите на култивирани миши сензорни неврони. J Physiol. 2006;577:815 28. doi: 10.1113/jphysiol.2006.117648. [PMC безплатна статия]�[PubMed]�[Крос Реф]
69. Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. Радиалното разтягане разкрива различни популации от механочувствителни соматосензорни неврони на бозайници.Proc Natl Acad Sci US A. 2008;105:20015 20. doi: 10.1073/pnas.0810801105. [PMC безплатна статия]�[PubMed]�[Крос Реф]
70. Crawford AC, Evans MG, Fettiplace R. Активиране и адаптиране на токове на трансдюсер в клетките на костенурката. J Physiol. 1989;419:405 34. [PMC безплатна статия]�[PubMed]
71. Ricci AJ, Wu YC, Fettiplace R. Ендогенният калциев буфер и времевият ход на адаптацията на трансдюсера в слуховите космени клетки.J Neurosci. 1998;18:8261 77. [PubMed]
72. Vollrath MA, Kwan KY, Corey DP. Микромашината на механотрансдукцията в космените клетки. Annu Rev Neurosci. 2007;30:339 65. doi: 10.1146/annurev.neuro.29.051605.112917. [Преглед] [PMC безплатна статия]�[PubMed]�[Крос Реф]
73. Goodman MB, Schwarz EM. Преобразуващо докосване в Caenorhabditis elegans. Annu Rev Physiol. 2003;65:429. doi: 52/annurev.physiol.10.1146. [Преглед] [PubMed]�[Крос Реф]
74. Waldmann R, Lazdunski MH. H(+)-зависими катионни канали: невронални киселинни сензори в семейството на NaC/DEG йонни канали. Curr Opin Neurobiol. 1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed]�[Крос Реф]
75. Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ и др. Йонният канал ASIC1 допринася за висцералната, но не и кожната механорецепторна функцияГастроентерология.2004;127:1739 47. doi: 10.1053/j.gastro.2004.08.061. [PubMed]�[Крос Реф]
76. Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE и др. Катионният канал DRASIC допринася за откриването на кожни докосвания и киселинни стимули при мишки.Неврон. 2001;32:1071 83. doi: 10.1016/S0896-6273(01)00547-5. [Erratum in: Neuron 2002 Jul 18;35] [2] [PubMed]�[Крос Реф]
77. Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M и др. Нокаутът на ASIC2 канала при мишки не нарушава кожната механосензация, висцералната механоноцицепция и слуха.J Physiol. 2004;558:659 69. doi: 10.1113/jphysiol.2004.066001. [PMC безплатна статия]�[PubMed]�[Крос Реф]
78. Damann N, Voets T, Nilius B. TRPs в нашите сетива. Curr Biol. 2008;18:R880 9. doi: 10.1016/j.cub.2008.07.063. [Преглед] [PubMed]�[Крос Реф]
79. Кристенсен AP, Кори DP. TRP канали в механочувствителност: директно или индиректно активиране? Nat Rev Neurosci. 2007;8:510 21. doi: 10.1038/nrn2149. [Преглед] [PubMed]�[Крос Реф]
80. Liedtke W, Tobin DM, Bargmann CI, Friedman JM. TRPV4 от бозайници (VR-OAC) насочва поведенческите реакции към осмотични и механични стимули в Caenorhabditis elegans.Proc Natl Acad Sci US A. 2003;100(Допълнение 2): 14531 6. doi: 10.1073/pnas.2235619100. [PMC безплатна статия]�[PubMed]�[Крос Реф]
81. Suzuki M, Mizuno A, Kodaira K, Imai M. Нарушено усещане за натиск при мишки без TRPV4. J Biol Chem. 2003;278:22664 8. doi: 10.1074/jbc.M302561200. [PubMed]�[Крос Реф]
82. Liedtke W, Choe Y, Mart -Renom MA, Bell AM, Denis CS, Sali A и др. Осмотично активиран канал, свързан с ванилоиден рецептор (VR-OAC), кандидат осморецептор за гръбначни животни.Клетка. 2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
83. Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Преходният рецепторен потенциал ванилоид 4 е от съществено значение при невропатична болка, предизвикана от химиотерапия при плъхове.J Neurosci. 2004;24:4444 52. doi: 10.1523/JNEUROSCI.0242-04.2004. [Erratum в: J Neurosci. 2004 юни;24] [23][PubMed]�[Крос Реф]
84. Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J и др. TRPA1 медиира възпалителните действия на дразнители на околната среда и проалгетични агенти.Клетка. 2006;124:1269 82. doi: 10.1016/j.cell.2006.02.023. [PubMed]�[Крос Реф]
85. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ и др. TRPA1 допринася за студена, механична и химическа ноцицепция, но не е от съществено значение за трансдукцията на космените клетки.Неврон. 2006;50: 277. doi: 89/j.neuron.10.1016. [PubMed]�[Крос Реф]
86. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ и др. Piezo1 и Piezo2 са основни компоненти на различни механично активирани катионни канали. Наука. 2010;330:55 60. doi: 10.1126/science.1193270. [PMC безплатна статия]�[PubMed]�[Крос Реф]
87. Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS и др. Пиезо протеините са порообразуващи субединици на механично активирани канали. Природа. 2012;483: 176 81. doi: 10.1038/nature10812.[PMC безплатна статия]�[PubMed]�[Крос Реф]
88. Bae C, Sachs F, Gottlieb PA. Механочувствителният йонен канал Piezo1 се инхибира от пептида GsMTx4.Биохимия. 2011;50:6295 300. doi: 10.1021/bi200770q. [PMC безплатна статия]�[PubMed][Крос Реф]
89. Ким SE, Coste B, Chadha A, Cook B, Patapoutian A. Ролята на Drosophila Piezo в механичното възприятие. Природа. 2012;483:209 12. doi: 10.1038/nature10801. [PMC безплатна статия]�[PubMed][Крос Реф]
90. Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. Мутациите в механотрансдукционния протеин PIEZO1 са свързани с наследствена ксероцитоза.Кръв. 2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
91. Kawashima Y, G loc GS, Kurima K, Labay V, Lelli A, Asai Y и др. Механотрансдукцията в космените клетки на вътрешното ухо на мишката изисква трансмембранни гени, подобни на канал.J Clin Invest. 2011;121:4796 809. doi: 10.1172/JCI60405. [PMC безплатна статия]�[PubMed]�[Крос Реф]
92. Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouch ne J и др. TMC1, но не и TMC2, е отговорен за автозомно-рецесивно несиндромно увреждане на слуха в тунизийски семейства.Audiol Neurootol. 2008;13:213 8. doi: 10.1159/000115430. [PubMed]�[Крос Реф]
93. Манджи SS, Милър KA, Williams LH, Dahl HH. Идентифициране на три нови щама миши със загуба на слуха с мутации в гена Tmc1.съм Джей Патол.2012;180:1560 9. doi: 10.1016/j.ajpath.2011.12.034.[PubMed]�[Крос Реф]
94. Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A и др. Протеин от стоматинов домейн, от съществено значение за усещането за докосване на мишката. Природа. 2007;445:206 9. doi: 10.1038/nature05394. [PubMed][Крос Реф]
95. Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, et al. Механотрансдукция на стоматин и сензорни неврони. J Неврофизиол. 2007;98:3802 8. doi: 10.1152/jn.00860.2007.[PubMed]�[Крос Реф]
96. Huang M, Gu G, Ferguson EL, Chalfie M. Стоматин-подобен протеин, необходим за механосензация в C. elegans. Природа. 1995;378:292�5. doi: 10.1038/378292a0.�[PubMed]�[Крос Реф]
97. Ху Дж., Чанг Л.Й., Кох М., Люин Г.Р. Доказателство за протеинова връзка, участваща в соматичното докосване. EMBO J. 2010;29:855 67. doi: 10.1038/emboj.2009.398. [PMC безплатна статия]�[PubMed]�[Крос Реф]
98. Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L, et al. Ламинин-332 координира механотрансдукцията и бифуркацията на растежния конус в сензорните неврони.Nat Neurosci.2011;14: 993 × 1000. doi: 10.1038/nn.2873. [PubMed]�[Крос Реф]
99. Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G и др. TWIK-1, вездесъщ човешки слабо навътре ректифициращ K+ канал с нова структура.EMBO J. 1996;15:1004 11.[PMC безплатна статия]�[PubMed]
100. Lesage F. Фармакология на невронните фонови калиеви канали. Неврофармакология.2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed]�[Крос Реф]
101. Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE и др. Анализ на разпределението на човешки два домена на пори калиеви канали в тъканите на централната нервна система и периферията.Brain Res Mol Brain Res. 2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Крос Реф]
102. Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor E. Механо- или киселинна стимулация, два интерактивни режима на активиране на TREK-1 калиевия канал. J Biol Chem. 1999;274:26691 6. doi: 10.1074/jbc.274.38.26691. [PubMed]�[Крос Реф]
103. Maingret F, Fosset M, Lesage F, Lazdunski M, Honor E. TRAAK е невронален механо-зависим K+ канал на бозайници. J Biol Chem. 1999;274:1381 7. doi: 10.1074/jbc.274.3.1381. [PubMed][Крос Реф]
104. Alloui A, Zimmermann K, Mamet J, Duprat F, Noïl J, Chemin J и др. TREK-1, K+ канал, участващ в полимодалното възприемане на болката. EMBO J. 2006;25:2368 76. doi: 10.1038/sj.emboj.7601116.[PMC безплатна статия]�[PubMed]�[Крос Реф]
105. No'l J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S и др. Механично активираните K+ канали TRAAK и TREK-1 контролират възприятието както за топло, така и за студено.EMBO J. 2009;28:1308 18. doi: 10.1038/emboj.2009.57. [PMC безплатна статия]�[PubMed]�[Крос Реф]
106. Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. K+ каналите на TRESK с две пори представляват значителен компонент на фоновите калиеви токове в ганглийните неврони на гръбначния корен на миши.J Physiol. 2007;585:867 79. doi: 10.1113/jphysiol.2007.145649.[PMC безплатна статия]�[PubMed]�[Крос Реф]
107. Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR и др. Остри агенти от сечуански пипер възбуждат сензорните неврони, като инхибират двупоровите калиеви канали.Nat Neurosci.2008;11: 772 × 9. doi: 10.1038/nn.2143. [PMC безплатна статия]�[PubMed]�[Крос Реф]
108. Lennertz RC, Tsunozaki M, Bautista DM, Stucky CL. Физиологична основа на изтръпващата парестезия, предизвикана от хидрокси-алфа-саншул.J Neurosci. 2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[PMC безплатна статия]�[PubMed]�[Крос Реф]
109. Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM и др. KCNQ4 K(+) каналите настройват механорецепторите за нормално докосване при мишка и човек. Nat Neurosci.2012;15: 138 × 45. doi: 10.1038/nn.2985. [PubMed]�[Крос Реф]
110. Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG и др. Генетична основа за механосензорни черти при хората. PLoS Biol. 2012;10:e1001318. doi: 10.1371/journal.pbio.1001318.[PMC безплатна статия]�[PubMed]�[Крос Реф]
111. Delmas P, Hao J, Rodat-Despoix L. Молекулярни механизми на механотрансдукция в сензорни неврони на бозайници. Nat Rev Neurosci. 2011;12: 139. doi: 53/nrn10.1038. [PubMed]�[Крос Реф]
Механорецептивна болка: Според CDC, повече от 50% от възрастните в САЩ (125 милиона) са имали мускулно-скелетни разстройство на болката през 2012 г.
Повече от 40% от възрастните с мускулно-скелетна болка са използвали допълнителен здравен подход по каквато и да е причина през 2012 г. Това е значително по-високо от употребата при хора без мускулно-скелетно заболяване (24.1%). Използването на допълнителни здравни подходи по каквато и да е причина сред хората с болки в шията или проблеми е повече от два пъти по-високо от употребата при хора без тези проблеми.
„Сред възрастните с разстройство на мускулно-скелетната болка, използването на какъвто и да е допълнителен здравен подход е най-високо сред тези с болки във врата или проблеми (50.6%), следвани от хора с други мускулно-скелетни проблеми (46.2%).
Употребата на допълнителни здравни подходи по каквато и да е причина сред хора с болки в шията или проблеми е повече от два пъти по-висока от употребата при хора без тези проблеми.
Механорецепторите са сензорни рецептори, които реагират на механичен натиск или изкривяване.
Те включват кожни рецептори за докосване, рецептори, които следят дължината и напрежението на мускулите, слухови и вестибуларни рецептори и други.
ТЕОРИЯ ЗА КОНТРОЛ НА БОЛКАТА
Безболезненото въвеждане затваря вратите за болезнено въвеждане.
Това предотвратява преминаването на болковите усещания до по-високи кортикални нива
Аферентите с малък диаметър (болка) възпрепятстват инхибирането на болката
Аферентите с голям диаметър (вибрация) са склонни да възбуждат инхибиране на болката.
Тази теория твърди, че не-ноцицептивните влакна могат да пречат на сигналите от болковите влакна, като следователно инхибират болката.
А с голям диаметър? влакната са неноцицептивни (не предават болкови стимули) и инхибират ефектите на изстрелване от A? и C влакна.
ДОРСАЛНА КОЛОНА МЕДИАЛНА ЛЕМНИСКА ПЪТЯ
ИЗПОЛЗВАНЕ НА ПЕРИФЕРНИ МЕХАНОРЕЦЕПТОРИ ЗА ПРОМЕНЯНЕ НА ВЪЗПРИЯТИЕТО НА БОЛКАТА
КАК МОЖЕМ ДА ПОМОГНЕМ?
РЕФЛЕКС НА ОТТЕГЛЯНЕ
Възбуден аферентен неврон стимулира възбуждащите интерневрони, които от своя страна стимулират еферентните моторни неврони, захранващи бицепса, мускула на ръката, който огъва (сгъва) лакътната става. Свиването на бицепса издърпва ръката от горещата печка.
Аферентният неврон също така стимулира инхибиторните интерневрони, които от своя страна инхибират еферентните неврони, захранващи трицепса, за да предотвратят свиването му. Този тип невронна връзка, включваща стимулиране на нервното снабдяване на един мускул и едновременно инхибиране на нервите към неговия антагонистичен мускул, е известно като реципрочно инхибиране.
Аферентният неврон все още стимулира други интерневрони, които пренасят сигнала нагоре по гръбначния мозък към мозъка чрез възходящ път. Едва когато импулсът достигне до сензорната област на кората, човекът осъзнава болката, нейното местоположение и вида на стимула. Освен това, когато импулсът достигне до мозъка, информацията може да се съхранява като памет и човекът може да мисли за случилото се.
РЕЦЕПТОРНА ТЕРАПИЯ
Корекции
Активирането на ставните механорецептори чрез хиропрактика може да модулира и „засенчи“ възприятието на мозъка за влакна с по-малък диаметър.
Повторението на активиране на ставните механорецептори може да създаде положителна пластичност в аферентните пътища.
Положителната пластичност може да изключи болката
Вибрация
Вибрационната стимулация при определени честоти може да промени възприятието за болка
Повторението на активиране на дисковете на Меркел и телцата на Мейснер може да създаде положителна пластичност в аферентните пътища.
Отново, положителната пластичност може да изключи болката
VIBRATION
Този тип устройство прилага синусоидални вибрации и предлага непрекъсната избираема амплитуда от 0-5.2mm в зависимост от позицията на крака и избираема честота от 5-30Hz.
Обучението по WBV изглежда е ефективна, безопасна и подходяща интервенция за седнали работещи служители с хронична болка в кръста.
Хомотопната вибро-тактилна стимулация доведе до 40% намаляване на болката от топлина във всички субектни групи. Изглежда, че разсейването не е повлияло на експерименталните оценки на болката.
Вибро-тактилната стимулация ефективно включва аналгетични механизми не само при NC, но и при пациенти с хронична мускулно-скелетна болка, включително FM.
ЛЕКО ДОКОСВАНЕ
Общо 44 здрави доброволци са изпитали топлинна болка и CT оптимални (бавно четкане) и CT неоптимални (бързо четкане или вибрации) стимули. Използвани са три различни експериментални парадигми: Едновременно приложение на топлинна болка и тактилна (бавно четкане или вибрация) стимулация; Бавно четкане, прилагано за променлива продължителност и интервали, предшестващо топлинна болка; Бавно срещу бързо четкане, предшестващо болка от топлина.
При хората основните мозъчни области, получаващи информация за C-LTMR, принадлежат към соматосензорната система и влияят на обработващите мозъчни мрежи като контралатералната задна островна кора или медиалната префронтална кора. Интензитетът на CT насоченото докосване се кодира в първичната и вторичната соматосензорна кора (S1 контралатерално, S2 двустранно), докато приятността е кодирана в предния цингулатен кортекс. C-LTMR също активират региони, участващи в обработката на възнаграждението (путамен и орбитофронтален кортекс) и в обработката на социални стимули (задна горна темпорална бразда).
ВСИЧКО ПЕРИФЕРНО ИМА ЦЕНТРАЛНО ПОСЛЕДСТВИЕ
КАЗУС
47-годишен мъж претърпя лява CVA през октомври 2017 г.
Не е движил дясната страна на тялото си след инцидента.
Представен в нашата клиника, защото иска „да се върне към нея“.
АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД
Дизартрия
Променено възприятие за болка
Трудност с проста математика
Отпуснат на RUE и RLE
АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД
Пациентът нямаше движение, докато не започнахме да тестваме усещане и рефлекси:
АЛОДИНИЯ: Отнася се до централна сенсибилизация на болка (повишена реакция на невроните) след нормално неболезнена, често повтаряща се стимулация.
Алодинията може да доведе до задействане на болков отговор от стимули, които обикновено не провокират болка.
Температурата или физическите стимули могат да провокират алодиния, която може да се почувства като усещане за парене и често се появява след нараняване на мястото.
Alldynia е различна от хипералгезията, екстремна, преувеличена реакция към стимул, който обикновено е болезнен.
ТЕРАПЕВТИЧНИ ИНТЕРВЕНЦИИ
Вибрация
Леко докосване
Акупресура
Акустични честоти
Корекции!
СЛЕД ДВА ДНИ
Механорецептивна болка и рецепторно базирана терапия
Защо локализираното увреждане или нараняване, причинено от травма, води до хронична, непреодолима болка при определени пациенти? Какво отговаря за превеждането на локално увреждане с остра болка в състояние на хронична болка? Защо някои болки реагират на противовъзпалителни лекарства и/или лекарства, докато други форми на болка изискват опиати?
Болка е сложен процес, включващ както периферната нервна система (ПНС), така и централната нервна система (ЦНС). Увреждането на тъканите задейства PNS, който предава сигнали през гръбначния мозък в мозъка, в който възниква усещане за болка. Какво обаче кара интензивното изживяване на болката да се превърне в непрестанен феномен? Може ли да се направи нещо, за да се предотврати? Доказателствата сочат, че хронична болка резултат от комбинация от механизми, като неврологични „спомени“ за предшестваща болка.
Ноцицепция: Най-простият път
Острата или ноцицептивна болка се характеризира като редовно изпитване на дискомфорт, което се появява в отговор на съвсем елементарно увреждане или нараняване. Той е защитен, предупреждава ни да се отдалечим от произхода на обидата и да се погрижим за травмата. Механизмите, които създават ноцицептивна болка, включват трансдукция, която разширява външната травматична стимулация в електрическа активност в специализирани ноцицептивни първични аферентни нерви. След това аферентните нерви провеждат сензорната информация от PNS към CNS.
В ЦНС данните за болката се предават от първичните сензорни неврони в централните проекционни клетки. След като информацията се прехвърли във всички онези области на мозъка, които са отговорни за нашето възприятие, се случва действителното сетивно преживяване. Ноцицептивната болка е сравнително проста реакция на особено прост, остър стимул. Но механиката, отговаряща за ноцицептивната болка, не може да идентифицира явления, като болка, която продължава въпреки отстраняването или излекуването на стимулацията, като например при фантомна болка в крайниците.
Болка и възпалителна реакция
При обстоятелства на по-тежко нараняване, като хирургични рани, увреждането на тъканите може да стимулира възпалителна реакция. Въпреки това, други състояния, особено артрит, също могат да се характеризират с продължаващи случаи на възпаление, свързани със симптоми на интензивна болка. Механизмите за този тип болка, свързана с увреждане на тъканите и възпалителен отговор, са различни от ноцицептивната болка с ранно предупреждение.
Наблюдавайки разреза или мястото на друго увреждане или нараняване, в нервната система възниква каскада от свръхвъзбудими събития. Това явление на телесното „навиване“ започва от кожата, където се потенцира по периферните нерви и завършва с реакция на свръхчувствителност по протежение на гръбначния мозък (дорзалния рог) и мозъка. След това възпалителните клетки обграждат областите на увреждане на тъканите и също така произвеждат цитокини и хемокини, вещества, които са предназначени да медиират процеса на заздравяване и регенерация на тъканите. Но тези агенти могат също да се считат за дразнители и да коригират свойствата на първичните сензорни неврони около зоната на травмата.
По този начин основните фактори, които предизвикват възпалителна болка, включват увреждане на високопраговите ноцицептори, известни като периферна сенсибилизация, промени и изменения на невроните в нервната система и усилване на възбудимостта на невроните в ЦНС. Това представлява централна сенсибилизация и е отговорно за свръхчувствителността, при която областите, съседни на тези на истинското нараняване, ще изпитват болка, сякаш са наранени. Тези тъкани могат също да реагират на стимулация, която обикновено не причинява болка, като докосване, носене на дрехи, лек натиск или дори разресване на собствената ви коса, сякаш са наистина болезнени, наричани алодиния.
Невропатичната болка е резултат от увреждане или нараняване на нервната система, като синдром на карпалния тунел, постхерпетична невралгия и диабетна невропатия. Въпреки че някои от механизмите, които изглежда причиняват невропатична болка, се припокриват с тези, отговорни за възпалителната болка, много от тях са различни и следователно ще се нуждаят от различен подход към тяхното управление.
Процесът на периферна и централна сенсибилизация се поддържа, поне теоретично и експериментално, по време на възбуждащия невротрансмитер, глутамат, за който се смята, че се освобождава, когато се активира N-метил-D-аспартат (NMDA) рецептор.
Нервната система се състои от инхибиторни или възбуждащи невротрансмитери. Повечето от това, което позволява на нашата нервна система да реагира по подходящ начин на увреждане или нараняване, е фината настройка или инхибиране на различни процеси. Превъзбуждането на нервната система се смята за проблем при редица различни заболявания. Например, свръхактивирането на NMDA рецептор може също да бъде свързано с афективни разстройства, симпатикови аномалии и дори толерантност към опиати.
Дори обикновената ноцицептивна болка до известна степен активира NMDA рецептора и се смята, че води до освобождаване на глутамат. Независимо от това, при невропатичната болка свръхчувствителността към NMDA рецептора е ключова.
При други видове хронична болка, като фибромиалгия и главоболие от тензионен тип, някои от механизмите, активни при възпалителна и невропатична болка, могат също да създадат подобни аномалии в болковата система, включително централна сенсибилизация, по-висока възбудимост на соматосензорните пътища и намаляване на инхибиторни механизми на централната нервна система.
Периферна сенсибилизация
Циклооксигеназата (COX) също играе важна функция както при периферната, така и при централната сенсибилизация. COX-2 е един от ензимите, които се индуцират по време на възпалителния процес; COX-2 превръща арахидоновата киселина в простагландини, които повишават чувствителността на периферните ноцицепторни терминали. На практика периферното възпаление също причинява производството на COX-2 от ЦНС. Сигналите от периферните ноцицептори са частично отговорни за това регулиране нагоре, но изглежда също така има хуморален компонент за трансдукцията на сигналите за болка през кръвно-мозъчната бариера.
Например, в експериментални модели, COX-2 се генерира от ЦНС, дори ако животните получат сензорен нервен блок преди периферна възпалителна стимулация. COX-2, който се експресира върху невроните на гръбначния рог на гръбначния мозък, освобождава простагландини, които действат върху централните терминали или пресинаптичните терминали на ноцицептивните сензорни влакна, за да увеличат освобождаването на предавателя. Освен това, те действат постсинаптично върху невроните на дорзалния рог, за да произведат директна деполяризация. И накрая, те инхибират активността на глициновия рецептор, а това е инхибиторен предавател. Следователно, простагландините създават повишаване на възбудимостта на централните неврони.
Мозъчна пластичност и централна сенсибилизация
Централната сенсибилизация описва промени, които се случват в мозъка в отговор на повтаряща се нервна стимулация. След повтарящи се стимули, количествата хормони и мозъчните електрически сигнали се променят, тъй като невроните развиват „памет“ за реагиране на тези признаци. Постоянната стимулация създава по-мощна мозъчна памет, така че мозъкът ще реагира по-бързо и ефективно, когато бъде подложен на същата стимулация в бъдеще. Последвалите модификации в мозъчното окабеляване и реакцията се наричат невронна пластичност, които описват способността на мозъка да се променя лесно или централна сенсибилизация. Следователно мозъкът се активира или сенсибилизира от предишни или повтарящи се стимули, за да стане по-възбудим.
Флуктуациите на централната сенсибилизация се появяват след многократни срещи с болка. Изследванията при животни показват, че многократното излагане на болезнена стимулация ще промени прага на болката на животното и ще доведе до по-силен отговор на болката. Изследователите смятат, че тези модификации могат да обяснят постоянната болка, която може да се появи дори след успешна операция на гърба. Въпреки че дисковата херния може да бъде отстранена от прищипан нерв, болката може да продължи като спомен за притискането на нерва. Новородените, подложени на обрязване без анестезия, ще реагират по-задълбочено на бъдеща болезнена стимулация, като рутинни инжекции, ваксинации и други болезнени процеси. Тези деца имат не само по-висока хемодинамична реакция, известна като тахикардия и тахипнея, но също така ще развият и засилен плач.
Тази неврологична памет на болката е изследвана подробно. В доклад за предишните си изследвания, Woolf отбелязва, че подобрената рефлексна възбудимост след увреждане или нараняване на периферната тъкан не разчита на непрекъснати периферни входни сигнали; по-скоро часове след периферна травма, рецептивните полета на невроните на гръбначния дорзален рог продължават да се увеличават. Изследователите също са документирали значението на гръбначния NMDA рецептор за индуцирането и поддържането на централна сенсибилизация.
Значение за управление на болката
След като централната сенсибилизация се установи, често са необходими по-големи дози аналгетици за нейното потискане. Превантивната аналгезия или терапията, преди болката да прогресира, може да намали ефектите на всички тези стимули върху ЦНС. Woolf демонстрира, че дозата морфин, необходима за спиране на централната свръхвъзбудимост, дадена преди кратка вредна електрическа стимулация при плъхове, е една десета от дозата, необходима за премахване на активността, след като тя е нараснала. Това се превежда в клиничната практика.
В клинично проучване на 60 пациенти, подложени на абдоминална хистеректомия, лица, които са получили 10 mg морфин интравенозно по време на въвеждане в анестезия, изискват значително по-малко морфин за контрол на следоперативната болка. Освен това чувствителността към болка около раната, наричана вторична хипералгезия, също е намалена в групата, третирана предварително с морфин. Превантивната аналгезия беше използвана със сравним успех в набор от хирургични настройки, включително преспинална операция и посттопедична операция.
Еднократна доза от 40 или 60 mg/kg ректален ацетаминофен има ясен морфин-спестяващ ефект при ежедневна хирургия при деца, ако се прилага при въвеждане на анестезия. Освен това, децата с достатъчна аналгезия с ацетаминофен изпитват значително по-малко постоперативно гадене и повръщане.
NMDA рецепторните антагонисти придават постоперативна аналгезия, когато се прилагат предоперативно. В литературата съществуват различни съобщения, подкрепящи употребата на кетамин и декстрометорфан в предоперативния период. При пациенти, подложени на реконструкция на преден кръстен лигамент, 24-часовата контролирана от пациента аналгезия на опиоидна консумация е значително по-малка в предоперативната категория декстрометорфан спрямо групата на плацебо.
В двойно-слепи, плацебо-контролирани проучвания, габапентин е показан като аналгетик за премедикация при пациенти, подложени на мастектомия и хистеректомия. Предоперативният перорален габапентин намалява резултатите от болката и постоперативната консумация на аналгетици без разлика в страничните ефекти в сравнение с плацебо.
Предоперативното приложение на нестероидни противовъзпалителни средства (НСПВС) показва значително намаляване на употребата на опиоиди след операцията. COX-2 са за предпочитане поради тяхната относителна липса на тромбоцитни ефекти и значителен стомашно-чревен профил на безопасност в сравнение с конвенционалните НСПВС. Целекоксиб, рофекоксиб, валдекоксиб и парекоксиб, приложени извън Съединените щати, предоперативно намаляват следоперативната употреба на наркотици с повече от 40 процента, като много пациенти използват по-малко от половината от опиоидите в сравнение с плацебо.
Блокирането на нервната проводимост в предоперативния период изглежда предотвратява развитието на централна сенсибилизация. Синдромът на фантомния крайник (PLS) се приписва на феномена на извиване на гръбначния стълб. Пациенти с ампутация
често се отстранява пареща или изтръпваща болка в частта на тялото. Една от възможните причини е, че нервните влакна в пъна се стимулират и мозъкът интерпретира сигналите като произхождащи от ампутираната част. Другото е пренареждането в рамките на кортикалните зони, така че тази област, да речем за ръката, сега реагира на сигнали от други части на тялото, но все пак ги интерпретира като идващи за ампутираната ръка.
Въпреки това, за пациенти, подложени на ампутация на долни крайници под епидурална анестезия, нито един от 11-те пациенти, получили лумбална епидурална блокада с бупивакаин и морфин за 72 часа преди операцията, не е развил PLS. За хора, подложени на обща анестезия без предходна лумбална епидурална блокада, 5 от 14 пациенти са имали PLS на 6 седмици, а 3 са продължили да изпитват PLS на 1 година.
Woolf и Chong отбелязват, че перфектното предоперативно, интраоперативно и следоперативно лечение се състои от „НСПВС за намаляване на активирането/централизацията на ноцицепторите, локални анестетици за блокиране на сензорния приток и централно действащи лекарства като опиати“. Намаляването на периоперативната болка с превантивни техники повишава удовлетвореността, ускорява изписването, щади употребата на опиоиди, заедно с намален запек, седация, гадене и задържане на урина и дори може да спре развитието на хронична болка. Анестезиолозите и хирурзите трябва да обмислят интегрирането на тези техники в ежедневните си практики.
Когато болката се появи в резултат на увреждане или нараняване в резултат на операция, гръбначният мозък може да достигне състояние на свръхвъзбудимост, при което се появяват прекомерни реакции на болка, които могат да продължат дни, седмици или дори години.
Защо локализираното нараняване в резултат на травма води до хронична, непреодолима болка при някои пациенти? Увреждането на тъканите води до съзвездие от промени във възбудимостта на гръбначния стълб, включително повишено спонтанно изстрелване, по-голяма амплитуда и дължина на реакцията, намален праг, засилено освобождаване от отговорност при повтаряща се стимулация и разширени рецептивни полета. Устойчивостта на тези промени, които общо се наричат централна сенсибилизация, изглежда е фундаментална за продължителното засилване на чувствителността към болката, която определя хроничната болка. Множество лекарства и/или лекарства, както и локална анестезия на нервната блокада могат да ограничат степента на спиране на централната нервна система (ЦНС), както се вижда от намалената болка и намалената консумация на опиоиди в моделите с превантивни аналгетици.
Прозрение на д-р Алекс Химензе
Хиропрактиката е алтернативна опция за лечение, която използва корекции на гръбначния стълб и ръчни манипулации за безопасно и ефективно възстановяване, както и поддържане на правилното подравняване на гръбначния стълб. Изследванията установиха, че изкривяването на гръбначния стълб или сублуксациите могат да доведат до хронична болка. Хиропрактиката обикновено се използва за управление на болката, дори ако симптомите не са свързани с нараняване и/или състояние в мускулно-скелетната и нервната система. Чрез внимателно повторно подравняване на гръбначния стълб, a хиропрактика може да помогне за намаляване на стреса и натиска от структурите около основния компонент на основата на тялото, като в крайна сметка осигурява облекчаване на болката.
Функция и болка на чревната нервна система
Когато става въпрос за намалена употреба на лекарства и/или лекарства, включително опиоиди, с цел предотвратяване на странични ефекти като стомашно-чревни здравословни проблеми, може да играе правилната функция на чревната нервна система.
Чревната нервна система (ENS) или вътрешната нервна система е един от ключовите клонове на автономната нервна система (ANS) и се състои от мрежеста система от нерви, която модулира ролята на стомашно-чревния тракт. Той е в състояние да действа независимо от симпатиковата и парасимпатиковата нервна система, въпреки че може да бъде засегната от тях. ENS може също да се нарече втори мозък. Произлиза от клетки на нервния гребен.
Човешката човешка нервна система се състои от около 500 милиона неврони, включително многобройните видове клетки на Догиел, приблизително една двеста от количеството неврони в мозъка. Ентеричната нервна система се вкарва в лигавицата на стомашно-чревната система, започвайки от хранопровода и се простира надолу до ануса. Клетките на Dogiel, известни също като клетки на Dogiel, се отнасят до някакъв вид мултиполярни надбъбречни тъкани в рамките на превертебралните симпатикови ганглии.
ENS е способна на автономни функции, като координация на рефлексите; въпреки че получава значителна инервация в автономната нервна система, тя работи и може да работи независимо от мозъка и гръбначния мозък. Ентеричната нервна система е описана като „втори мозък“ по редица причини. Чревната нервна система може да работи автономно. Обикновено комуникира с централната нервна система (ЦНС) чрез парасимпатиковата, или чрез блуждаещия нерв, и симпатиковата, тоест чрез превертебралните ганглии, нервни системи. Въпреки това, проучванията при гръбначни животни разкриват, че когато блуждаещият нерв е прекъснат, чревната нервна система продължава да функционира.
При гръбначните, чревната нервна система включва еферентни неврони, аферентни неврони и интерневрони, всички от които правят чревната нервна система способна да носи рефлекси и да действа като интегриращ център при липса на вход от ЦНС. Сетивните неврони съобщават за механични и химични условия. Чревната нервна система има способността да променя своя отговор въз основа на фактори като хранителен и обемен състав. В допълнение, ENS съдържа поддържащи клетки, които са много като астроглия на мозъка и дифузионна бариера около капилярите около ганглиите, която е като кръвно-мозъчната бариера на кръвоносните съдове.
Чревната нервна система (ENS) играе основна роля във възпалителните и ноцицептивните процеси. Лекарствата и/или лекарствата, които взаимодействат с ENS, наскоро предизвикаха значителен интерес поради способността им да регулират множество аспекти на чревната физиология и патофизиология. По-специално, експерименти с животни показват, че рецепторите, активирани от протеиназа (PARs) могат да бъдат от съществено значение за неврогенното възпаление в червата. Освен това, PAR2 агонистите изглежда предизвикват чревна свръхчувствителност и хипералгетични състояния, което предполага роля на този рецептор във възприемането на висцерална болка.
Освен това, PARs, заедно с протеиназите, които ги активират, представляват вълнуващи нови цели за терапевтична интервенция върху ENS. Обхватът на нашата информация е ограничен до хиропрактика, както и до наранявания и състояния на гръбначния стълб. За да обсъдите темата, моля не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителни теми: Сцинтика
Ишиас се нарича медицинско като събиране на симптоми, а не като единична травма и / или състояние. Симптомите на болката в седалищния нерв или ишиас може да варират по честота и интензивност, но най-често се описват като внезапна, остра (ножообразна) или електрическа болка, която излъчва от ниското гърба надолу до бедрата, бедрата, бедрата и краката в крака. Други симптоми на ишиас може да включват усещане за изтръпване или парене, изтръпване и слабост по дължината на седалищния нерв. Склерата най-често засяга индивидите между годините 30 и 50. Тя често може да се развие в резултат на дегенерация на гръбнака, дължаща се на възрастта, обаче, компресията и дразненето на седалищния нерв, причинени от изпъкнал или дискова херния, сред другите проблеми, свързани със здравето на гръбначния стълб, могат също да причинят болки в седалищния нерв.
Инструментът Find A Practitioner на IFM е най-голямата мрежа за препращане във функционалната медицина, създадена, за да помогне на пациентите да намерят практикуващи функционална медицина навсякъде по света. Сертифицираните практикуващи от IFM са изброени на първо място в резултатите от търсенето, като се има предвид тяхното обширно образование по функционална медицина

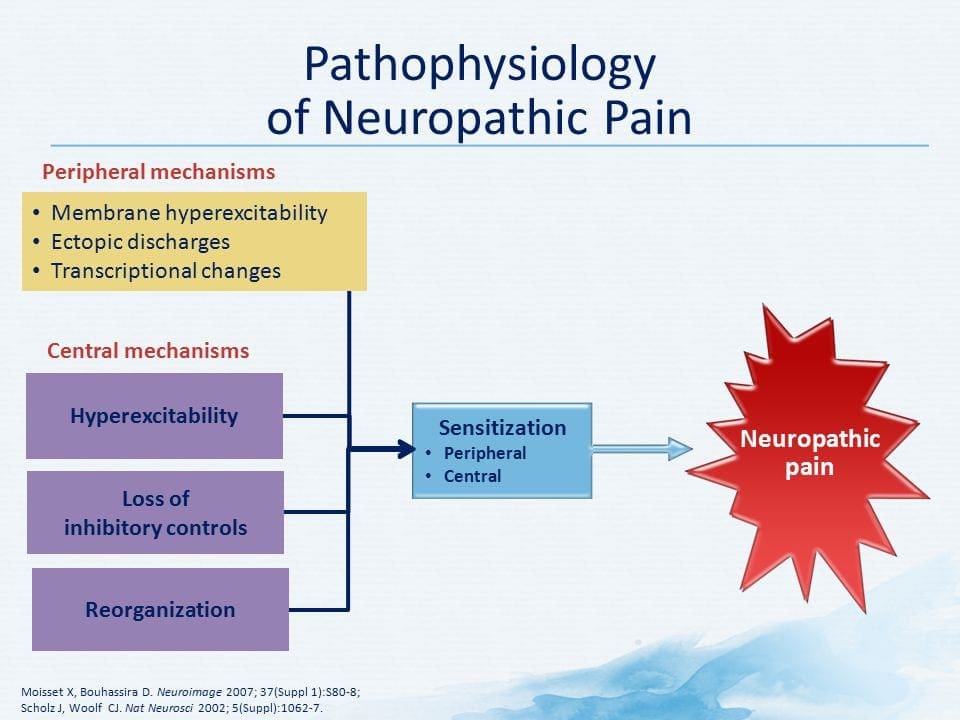
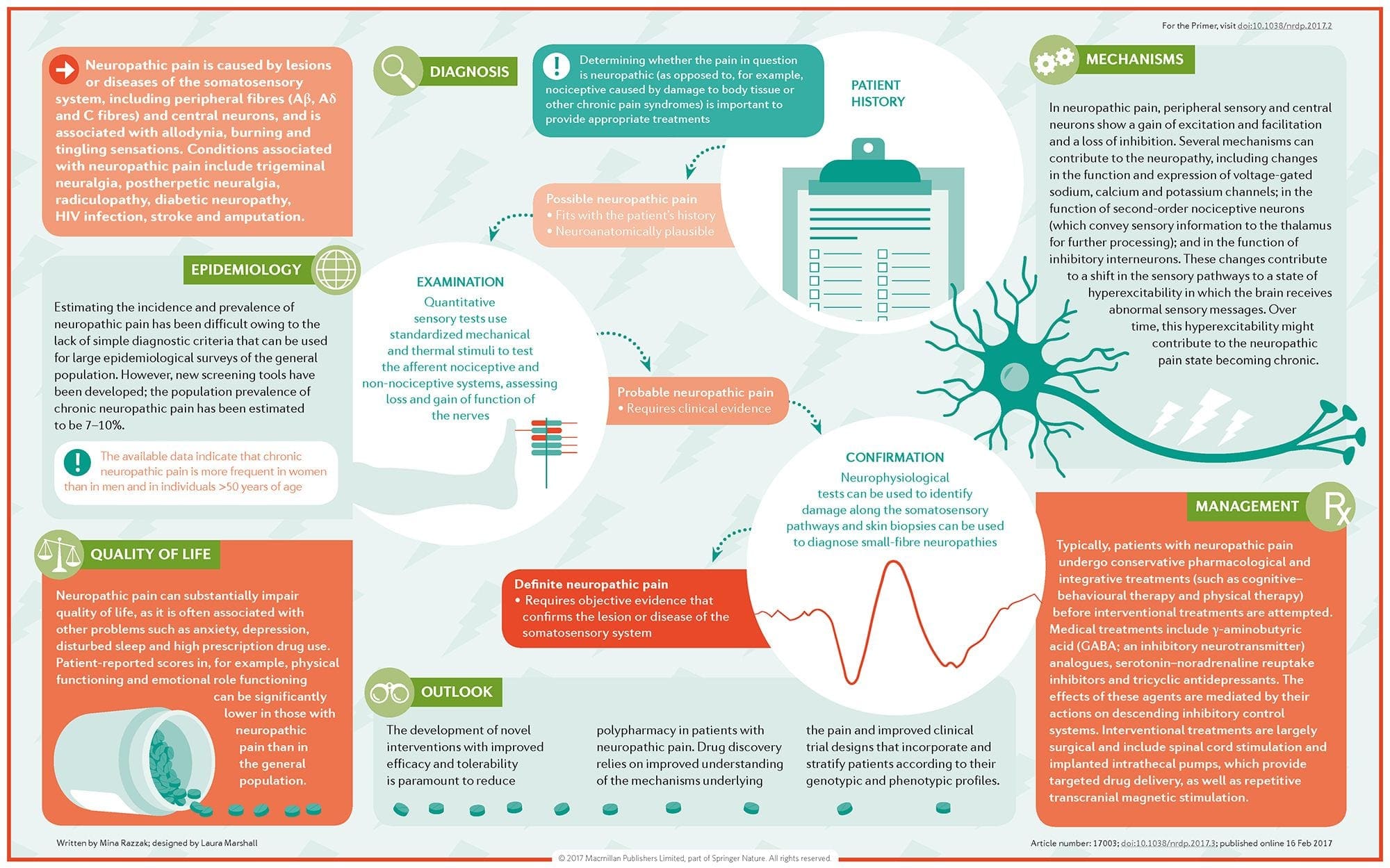




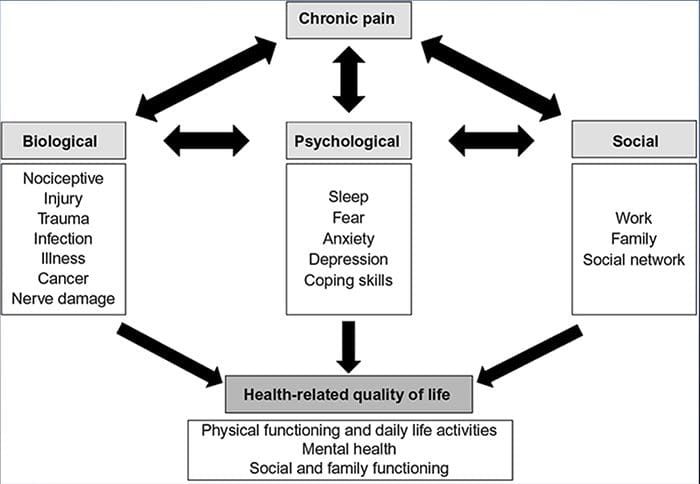
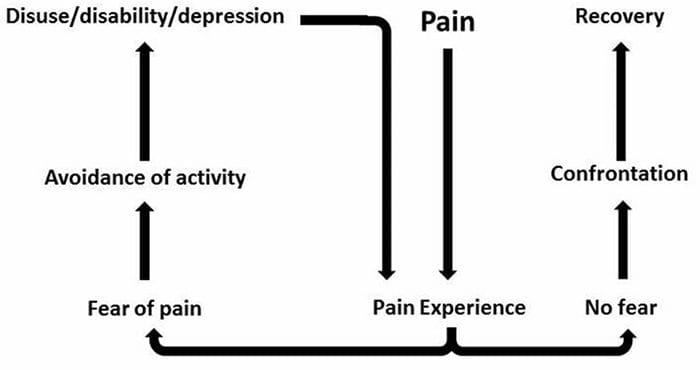
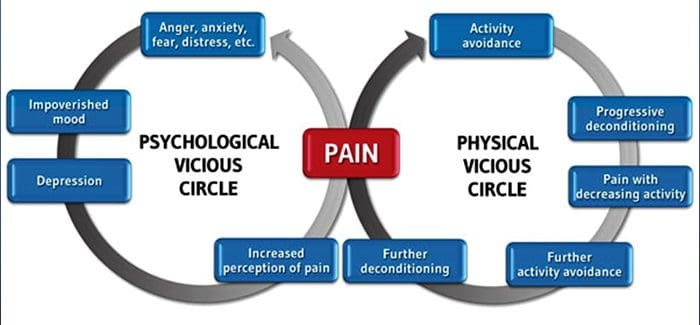
















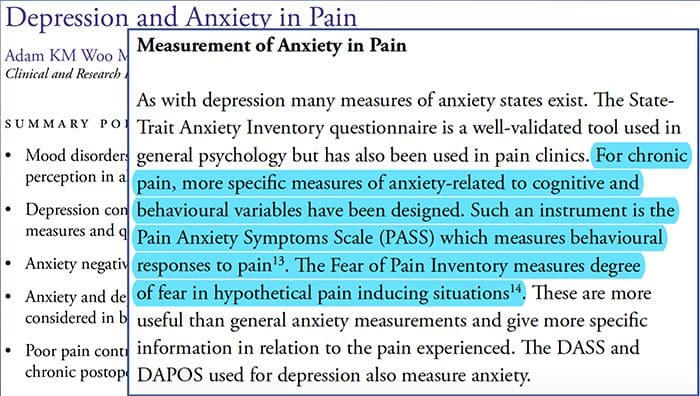


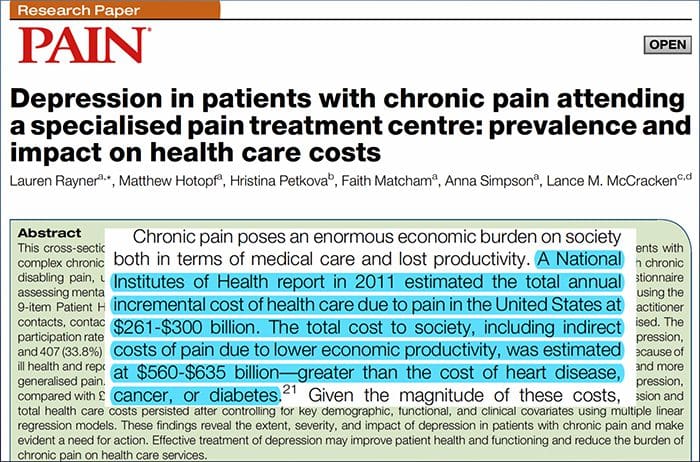



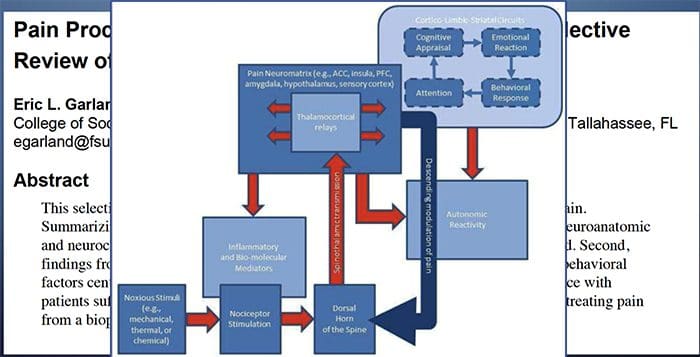






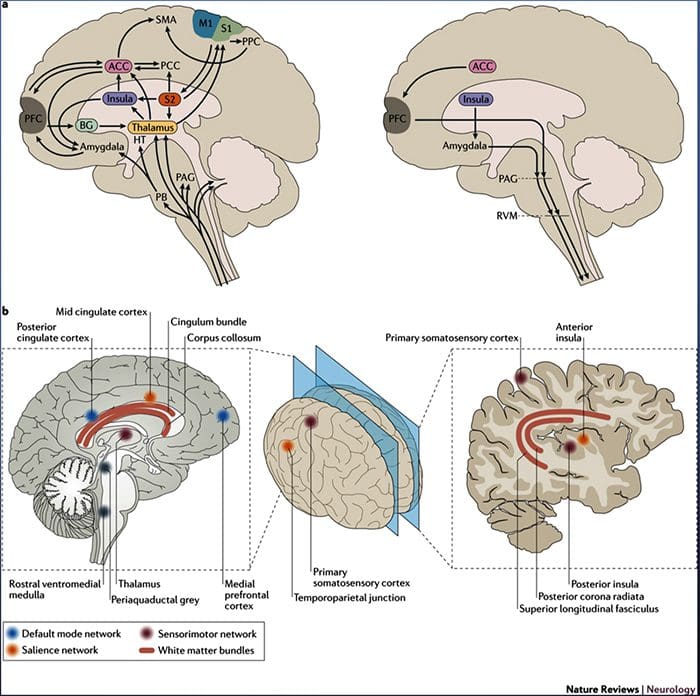

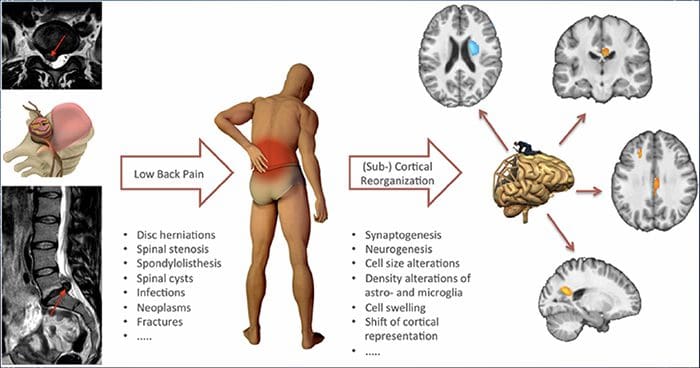

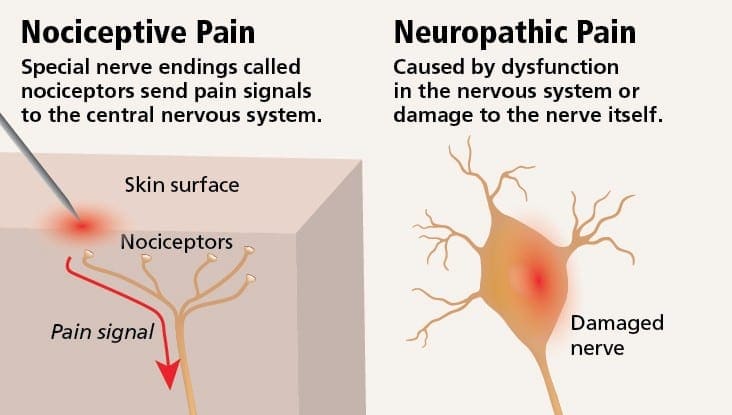
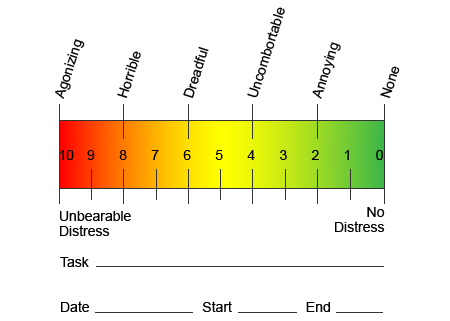


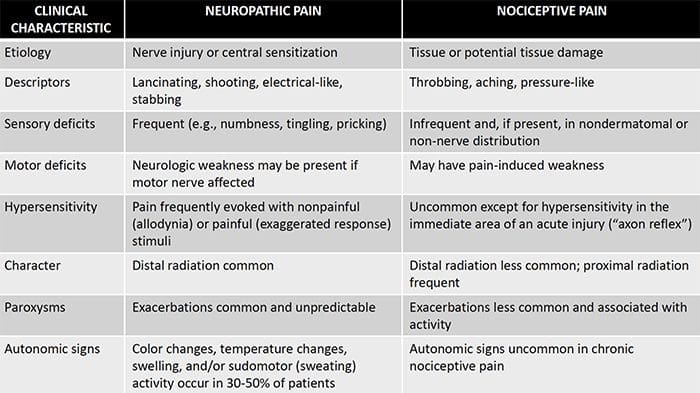
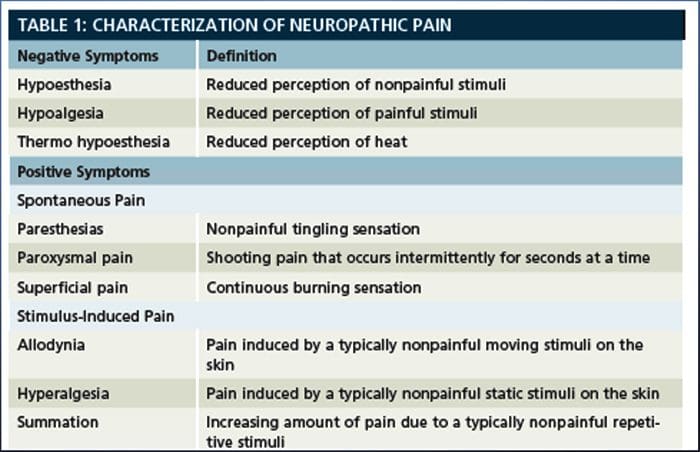
 ПАТОГЕНЕЗА НА НЕВРОПАТИЧНА БОЛКА
ПАТОГЕНЕЗА НА НЕВРОПАТИЧНА БОЛКА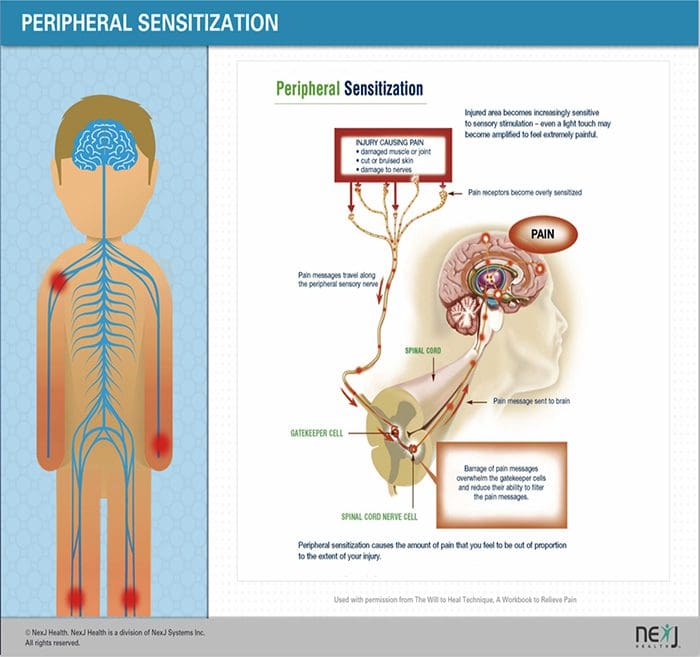
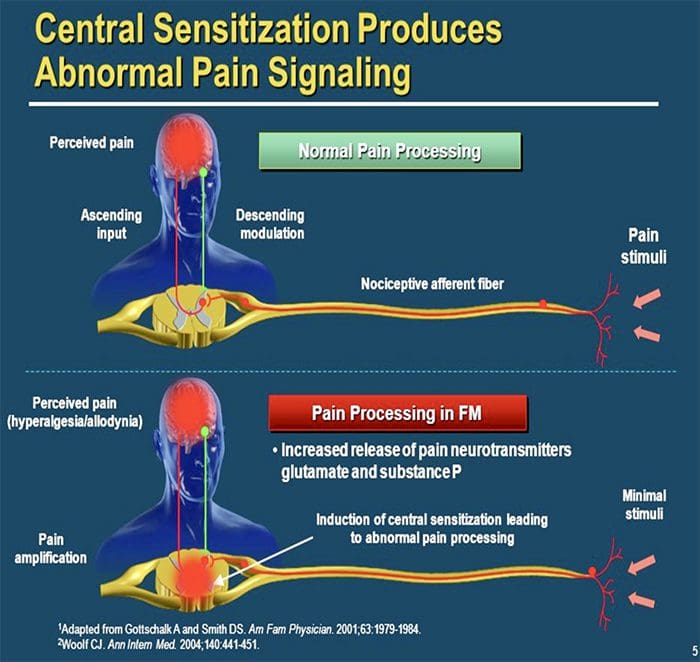
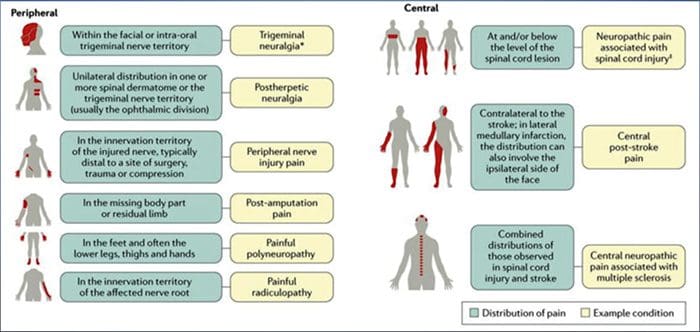 ЧЕСТИ ПРИЧИНИ
ЧЕСТИ ПРИЧИНИ
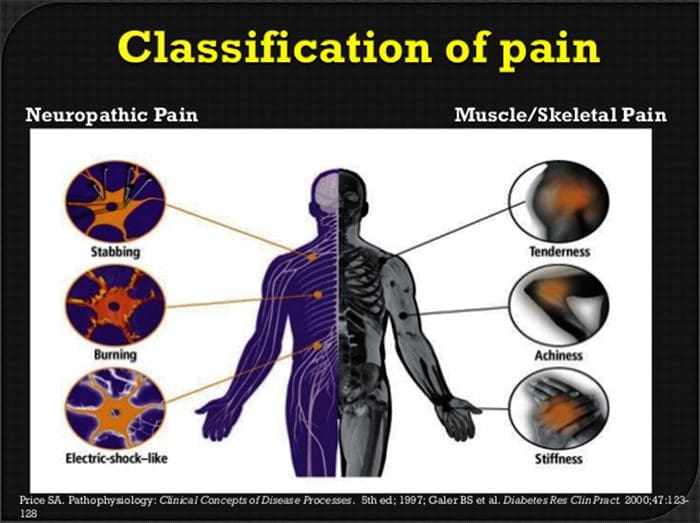
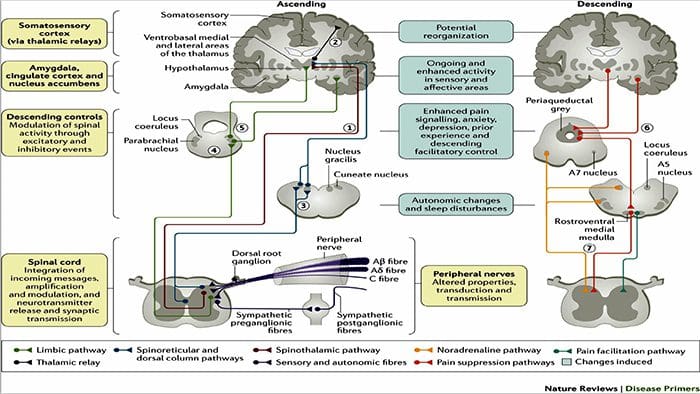
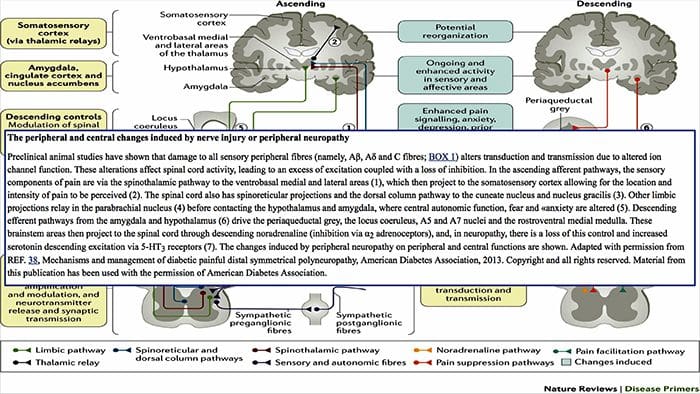

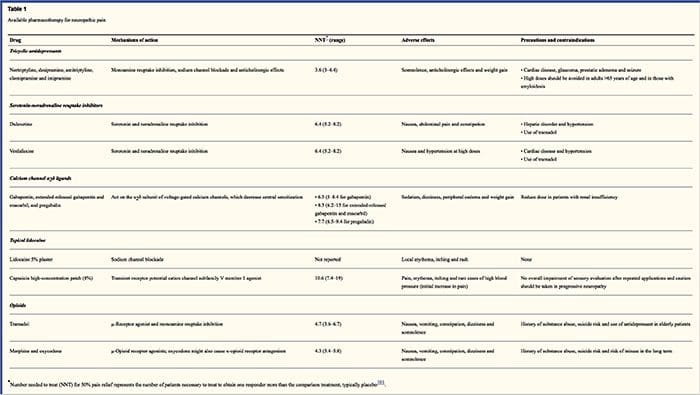



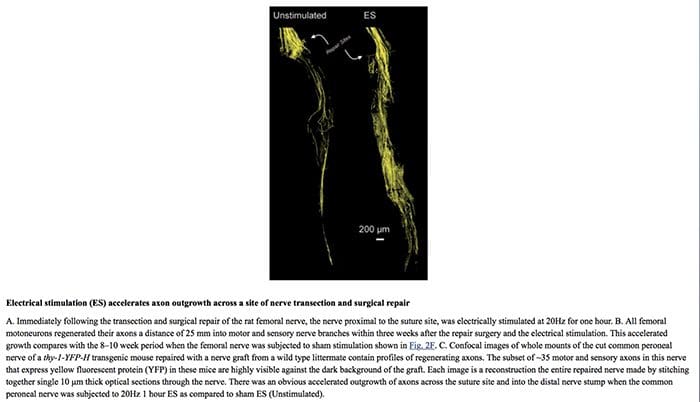

 ФАНТОМНА БОЛКА В КРАЙНИКАТА И РАЗОШЕНА РЕАЛНОСТ
ФАНТОМНА БОЛКА В КРАЙНИКАТА И РАЗОШЕНА РЕАЛНОСТ
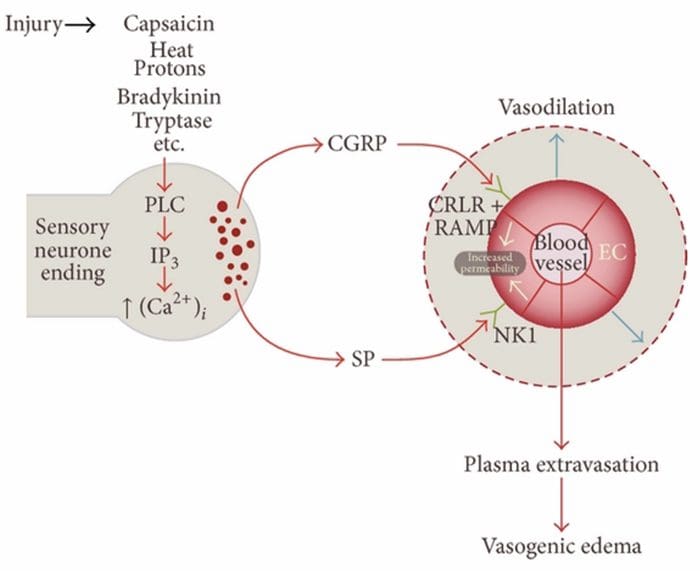 ЧЕСТИ ПРИЧИНИ
ЧЕСТИ ПРИЧИНИ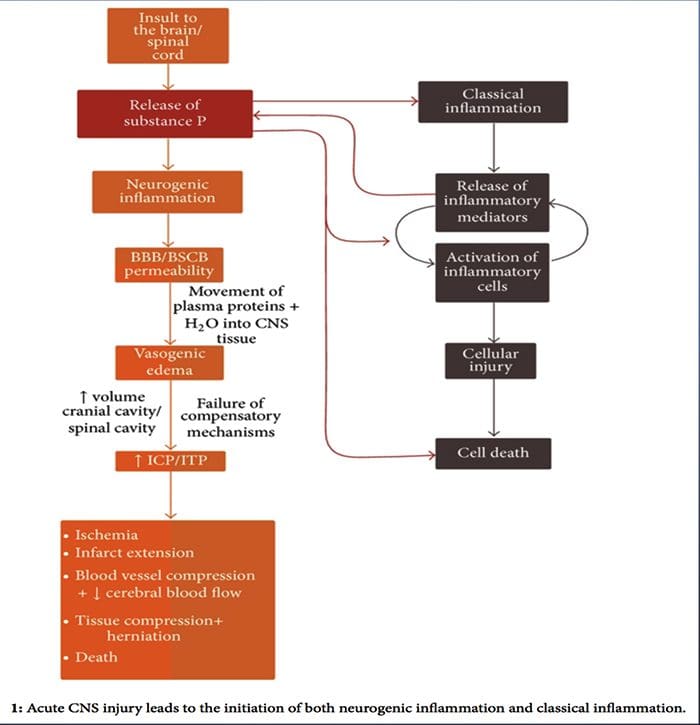


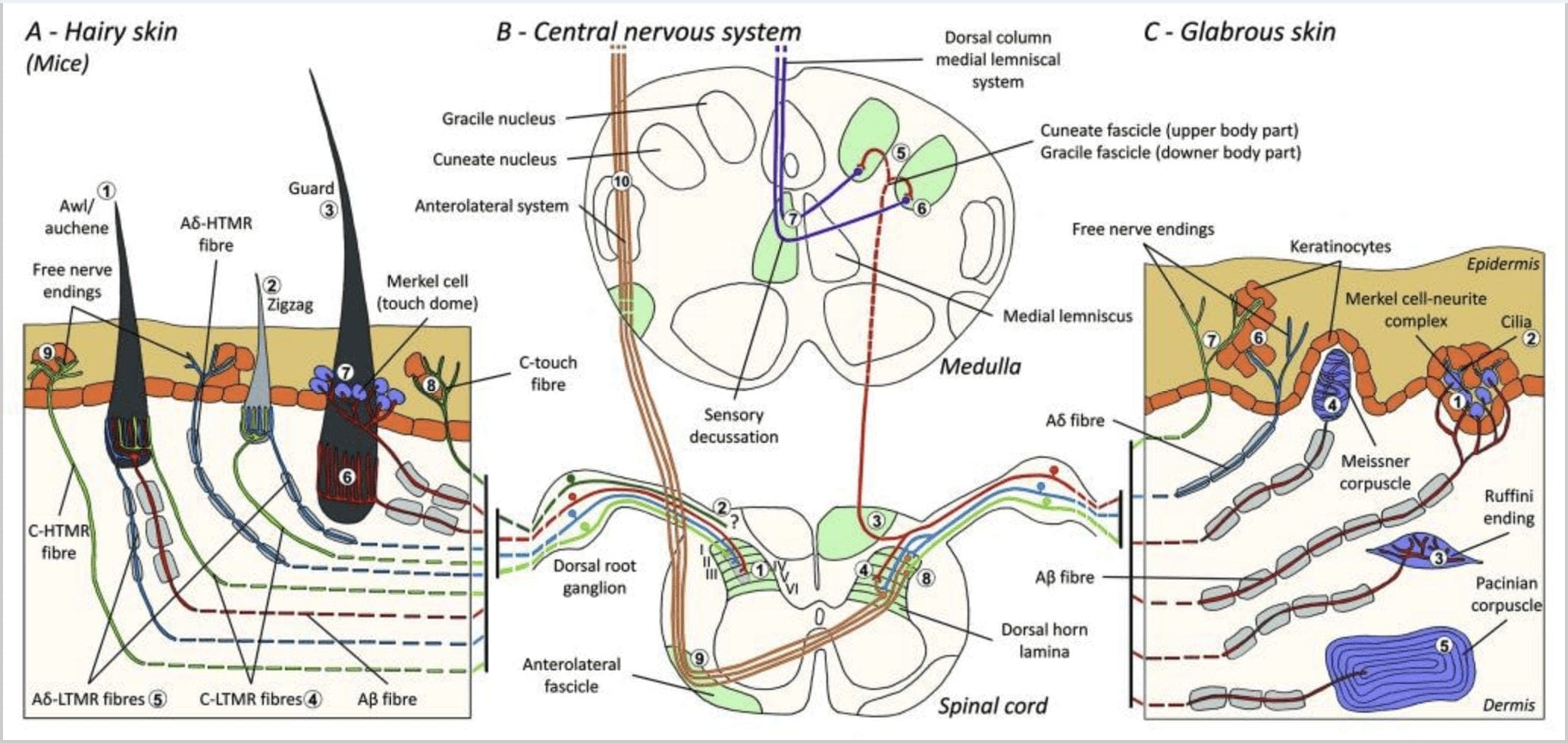
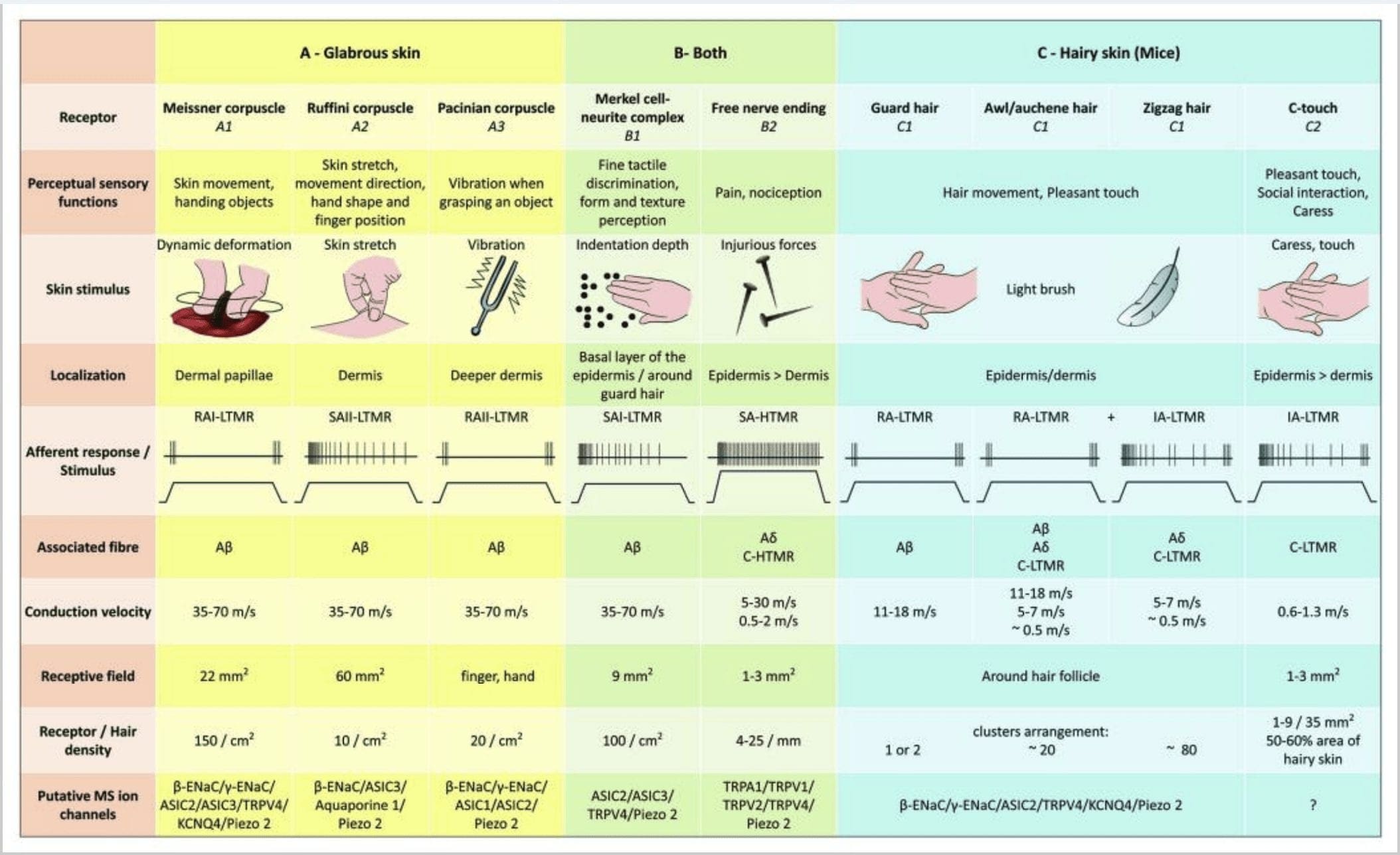

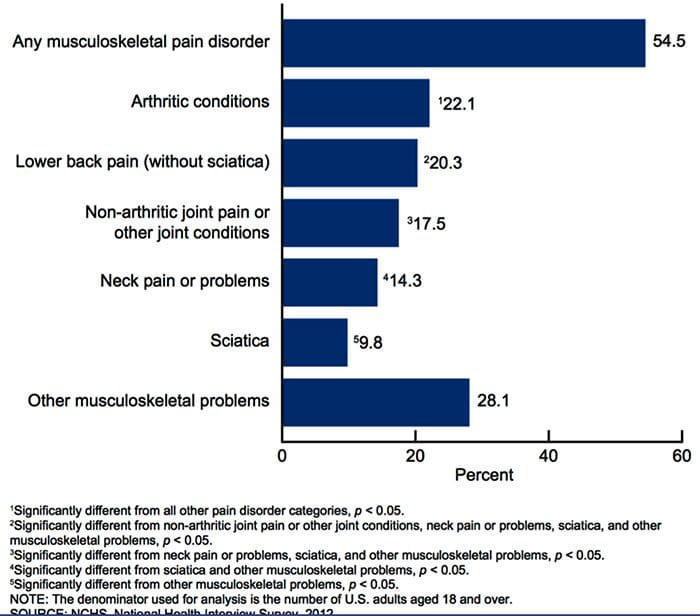
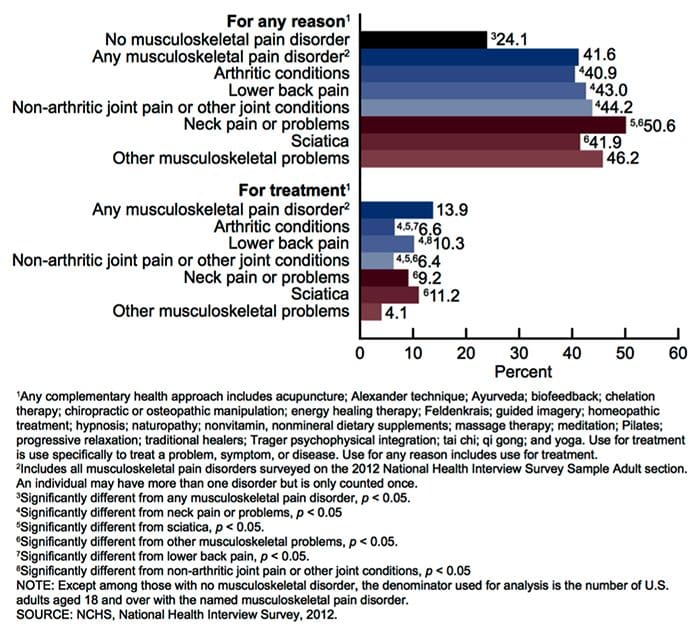
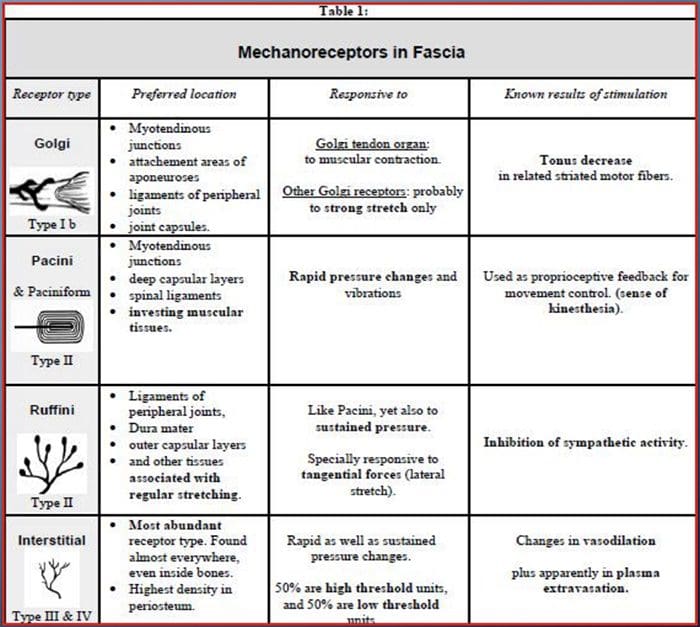 ТЕОРИЯ ЗА КОНТРОЛ НА БОЛКАТА
ТЕОРИЯ ЗА КОНТРОЛ НА БОЛКАТА
 ДОРСАЛНА КОЛОНА МЕДИАЛНА ЛЕМНИСКА ПЪТЯ
ДОРСАЛНА КОЛОНА МЕДИАЛНА ЛЕМНИСКА ПЪТЯ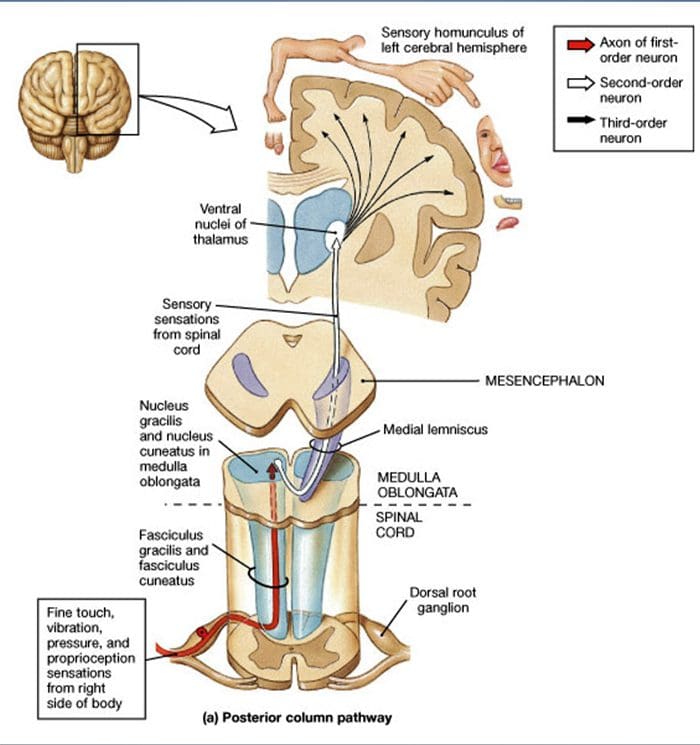 ИЗПОЛЗВАНЕ НА ПЕРИФЕРНИ МЕХАНОРЕЦЕПТОРИ ЗА ПРОМЕНЯНЕ НА ВЪЗПРИЯТИЕТО НА БОЛКАТА
ИЗПОЛЗВАНЕ НА ПЕРИФЕРНИ МЕХАНОРЕЦЕПТОРИ ЗА ПРОМЕНЯНЕ НА ВЪЗПРИЯТИЕТО НА БОЛКАТА
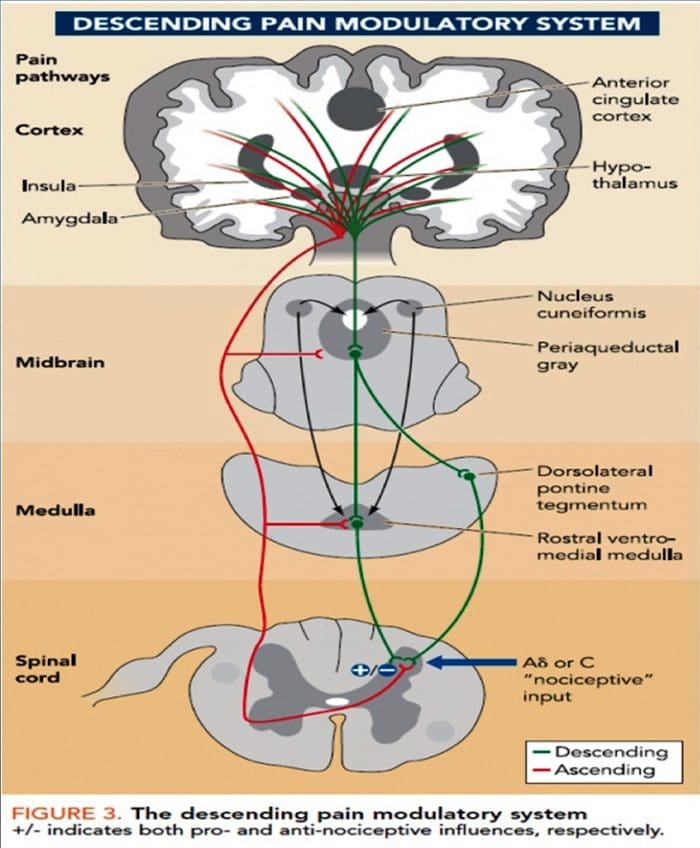
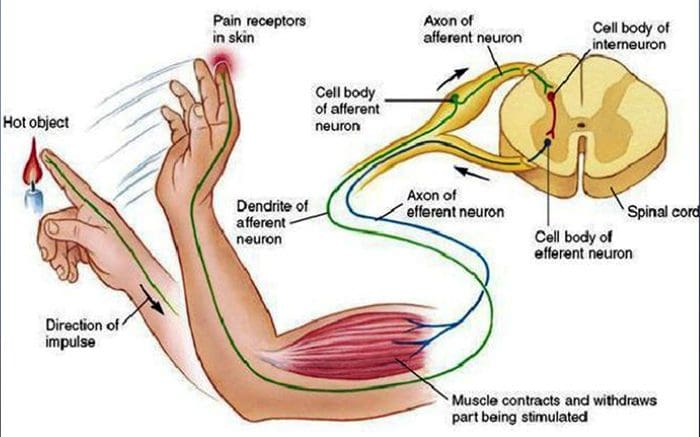
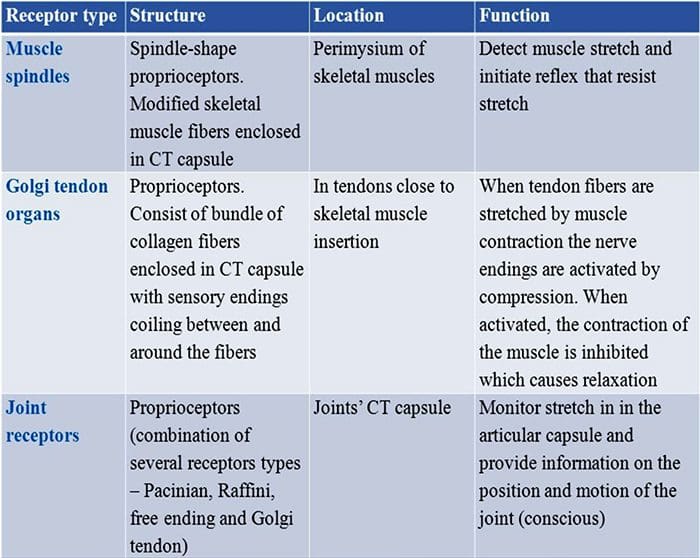
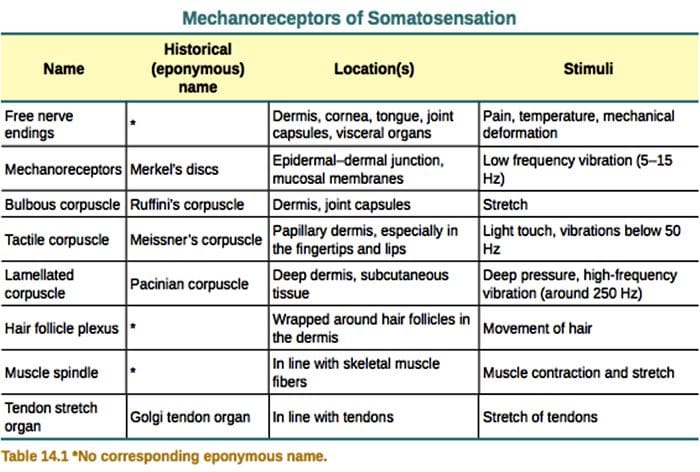 VIBRATION
VIBRATION
 ЛЕКО ДОКОСВАНЕ
ЛЕКО ДОКОСВАНЕ



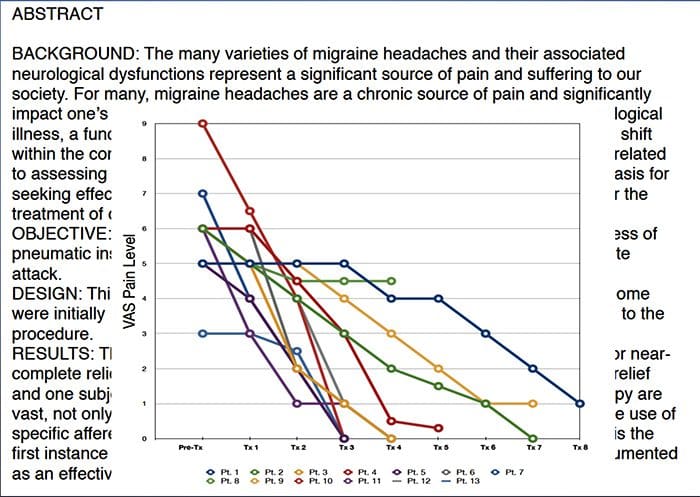


 КАЗУС
КАЗУС АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД
АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД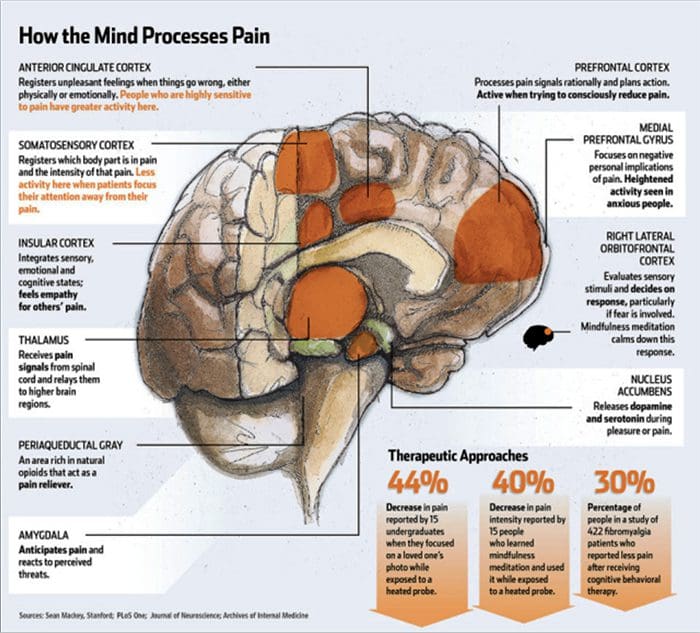 АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД
АКЦЕНТИ НА ФИЗИЧЕСКИ ИЗГЛЕД
 СЛЕД ДВА ДНИ
СЛЕД ДВА ДНИ