Back Clinic Екип по хиропрактика и функционална медицина против стареене. Тялото ни е в постоянна и безкрайна битка за оцеляване. Клетките се раждат, клетките се унищожават. Учените изчисляват, че всяка клетка трябва да издържи над 10,000 XNUMX индивидуални атаки от реактивни кислородни видове (ROS) или свободни радикали. Без провал, тялото има невероятна система за самолечение, която издържа на атаката и възстановява това, което е било повредено или унищожено. Това е красотата на нашия дизайн.
Да разбере биологията на стареенето и да преведе научното разбиране в интервенции, които подобряват здравето в края на живота чрез лечение. Полезно е да имате ясна, консенсусна гледна точка за това какво точно представлява лечението против стареене.
Още преди дните на търсенето на дълголетие на Понсе де Леон, човекът винаги е бил примамван от шанса за вечна младост. Хиропрактиката с нейното движение за здраве е мощен метод за стабилизиране и засилване на тази способност за самолечение. Д-р Алекс Хименес обсъжда концепциите около пандора против стареене.
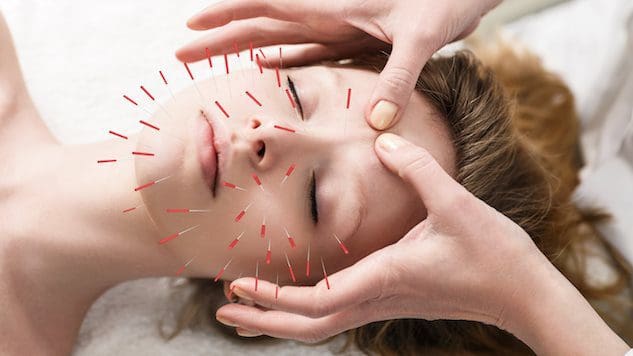
За хора, които искат да подобрят или поддържат здравето на кожата, може ли включването на акупунктура да помогне за подобряване на кожата и борба с процеса на стареене?
Козметична акупунктура
Козметичната акупунктура следва традиционната акупунктурна практика на поставяне на игла. Целта е да се обърнат признаците на стареене и да се подобри здравето на кожата. Понякога се нарича акупунктурно подмладяване на лицето, което се използва като алтернатива на хирургичния лифтинг на лицето и други конвенционални процедури. Предварителните проучвания изследват как може да помогне за премахване на старчески петна, повдигане на увиснали клепачи и намаляване на бръчките. (Younghee Yun и др., 2013 г)
Как действа акупунктурата
В традиционната китайска медицина или ТКМ, акупунктурата отдавна се използва за подобряване на потока на енергия – чи или чи – в тялото. Смята се, че тази енергия циркулира през енергийни пътища, известни като меридиани. Когато се появят здравословни проблеми, според ТКМ има препятствия или блокажи в кръвообращението.
Специалистите по акупунктура могат да възстановят оптималната циркулация/поток и да подобрят здравето чрез поставяне на игли в определени акупунктурни точки. (Национални институти по здравеопазване, 2007 г)
Козметична акупунктура
Твърди се, че козметичната акупунктура подобрява здравето на кожата и действа като лечение против стареене, като стимулира производството на колаген. Този протеин е основен компонент на кожата. Вътрешният слой на кожата губи колаген и стегнатост с напредване на възрастта на тялото. Необходими са обаче допълнителни изследвания в подкрепа на твърдението, че акупунктурата може да стимулира производството на колаген. Някои предполагат, че козметичната акупунктура помага за подмладяване на кожата чрез подобряване на цялостната енергия на тялото. Едно проучване установи, че хората виждат подобрения след пет сесии на лицева козметична акупунктура. (Younghee Yun и др., 2013 г) Въпреки това се препоръчва да се извършват десет процедури веднъж или два пъти седмично за оптимални резултати. След това се правят поддържащи лечения на всеки четири до осем седмици. За разлика от ботокса или дермалните филъри, козметичната акупунктура не е бързо решение. Фокусът е да се създадат дългосрочни промени в кожата и тялото, което означава подобряване на:
Когато иглите се вкарат в кожата, те създават рани, известни като положителни микротравми. Естествените лечебни и възстановителни способности на тялото се активират, когато усети тези рани. Тези пробиви стимулират лимфната и кръвоносната системи, които доставят хранителни вещества и кислород до клетките на кожата, подхранвайки ги отвътре навън.
Това помага за изравняване на тена и насърчава блясъка на кожата.
Положителните микротравми също стимулират производството на колаген.
Това спомага за подобряване на еластичността, минимизиране на линиите и бръчките.
Алтернативи
Няколко природни средства могат да помогнат за подобряване на здравето на кожата и да предложат ползи против стареене. Керамидите са мастна молекула, намираща се естествено в горния слой на кожата и съставка, използвана в продуктите за грижа за кожата. Те могат да предпазят от сухота на кожата, свързана със стареенето. (L Di Marzio 2008) Предварителните изследвания показват, че нанасянето на бял чай върху кожата може да се бори с разграждането на колаген и еластин – протеин, който поддържа еластичността на кожата и предотвратява увисването). Има също доказателства, че естествени вещества като арганово масло, масло от пореч и морски зърнастец могат да предложат овлажняващи ползи, които могат да подобрят кожата.(Tamsyn SA Thring et al., 2009)
Докато са необходими допълнителни доказателства за козметичната акупунктура, интегрирането на акупунктурата може да помогне за справяне със стреса и подобряване на цялостното здраве. Хората, обмислящи козметична акупунктура, трябва да се консултират с основния си доставчик на здравни услуги, за да видят дали е подходяща за тях.
Подобряване на здравето заедно: Възприемане на мултидисциплинарна оценка и лечение
Препратки
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS, & Choi, I. (2013). Ефект от козметичната акупунктура на лицето върху еластичността на лицето: отворено пилотно проучване с една ръка. Допълнителна и алтернативна медицина, базирана на доказателства: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Национален център за допълнителна и алтернативна медицина. (2007). Акупунктура: Въведение. Уебсайт на Националния център за допълнителна и алтернативна медицина. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH и Tsuji, R. (2021). Надеждност и валидност на лист за проверка на лицето (FCS): Контролен списък за самозадоволяване с козметична акупунктура. Лекарства (Базел, Швейцария), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Повишаване на нивата на кожния серамид при възрастни субекти след краткосрочно локално приложение на бактериална сфингомиелиназа от Streptococcus thermophilus. Международно списание за имунопатология и фармакология, 21 (1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009). Антиколагеназна, антиеластазна и антиоксидантна активност на екстракти от 21 растения. BMC допълваща и алтернативна медицина, 9, 27. doi.org/10.1186/1472-6882-9-27
Поддържането на гръбначния стълб на индивида в топ форма означава по-малко болка и повече мобилност, гъвкавост и свобода. Тялото се износва и е естествен ефект от стареенето, което се случва на всеки един от нас. Проблемите с гръбначния стълб, свързани със стареенето, могат да станат сериозни, ако не бъдат разгледани и приети с упражнения, разтягане и хиропрактика.
Стареене и гръб
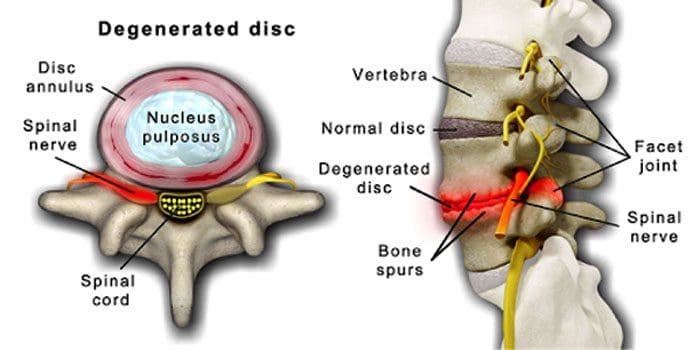
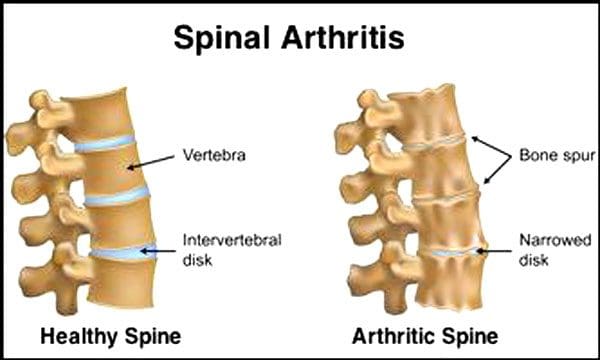
Нормално е гръбначните дискове и стави да се влошават с възрастта. Спиналната стеноза или стесняването на гръбначния канал също може да бъде част от процеса на стареене. Две условия, породени от стареенето, са дегенеративно дискова болест намлява артрит което също може да включва втвърдяване на гръбначните връзки и остеопороза.
Дегенеративна дискова болест се среща при 40% от хората на 40-годишна възраст
Увеличава се до 80% за лица на 80 и повече години.
То се центрира наоколо дискове, които постепенно се променят от предимно водни до предимно мазнини.
Когато е дебела, дисковете се стесняват и губят еластичност.
Това казват Центровете за контрол и превенция на заболяванията 23% от възрастните американци имат артрит. Това е състояние, което засяга главно фасетните стави. Ставите се подуват, което намалява обхвата на движение и може да засегне гръбначните нерви, причинявайки болка, слабост и ишиас. С времето връзките около и в гръбначния стълб се втвърдяват, намалявайки обхвата на движение, причинявайки стеноза. Загубата на костна маса или остеопорозата се причинява от промени в хормоните и други фактори като храненето. Стареенето е естествен процес, но хората могат да помогнат на гръбначния си стълб да останат в топ форма, независимо на колко години са.
Практикуване на здравословна стойка
Веднага правилна здравословна механика на тялото трябва. Да бъдеш наясно и да се съобразяваш с позата на тялото поддържа подравняване и поддържа тялото балансирано. Здравословната стойка ще помогне за намаляване на ефектите от:
Стеноза на гръбначния стълб
Дегенеративно дискова болест
херния
Риск от фрактури на гръбначния стълб
Практикуването на правилна стойка включва:
Намалете прегърбването
Уверете се, че работната станция е в най-добра форма и ергономична
Уверете се, че сте сгъвали коленете при повдигане и поддържайте гръбнака възможно най-вертикално.
Йога
Йога може да бъде много полезно за по-здрав и по-млад гръбначен стълб. Йога изпълнява три области за поддържане на гръбначния стълб в топ форма, Това включва:
Редовно упражнение
Поддържа гъвкавост
Постига идеално телесно тегло
Йогата е упражнение за гръбначния стълб, което се противопоставя на възрастта. Защото то:
Поддържа силата
Гъвкавост
Поза
Баланс
Може да бъде полезен при различни заболявания на гръбначния стълб, по-специално при артритна болка
Паданията могат да причинят сериозни наранявания. Йога също може да помогне за работата на баланса.
Вижте хиропрактик
Превантивната медицина е ключът към поддържането на тялото здраво, младо и възможно най-силно. Хиропрактикският преглед може да определи дали има проблеми с гръбначния стълб и да постави диагноза за разработване на оптимален план за лечение. Ако функцията на тялото е ограничена поради болка в гърба и/или краката, свържете се с Клиника по медицинска хиропрактика и функционална медицина за травми и върнете гръбначния стълб в най-добра форма.
телесния състав
Упражнение/Къдрици с топка за стабилност
Това упражнение работи мускулни групи, специфични за силата на гръбначния стълб и включва:
прасците
Глюте
Дълбок корем
Тазобедрени абдуктори и ротатори
Упражнения като това са един от най-ефективните начини за изграждане на функционална сила и издръжливост на подколенните сухожилия, бедрата и предотвратяване на наранявания. За да направите тази тренировка:
Легнете по гръб със свити колене
Повдигнете краката нагоре, така че долната част на стъпалата да лежи върху топката за упражнения
Разточете краката си, докато станат прави
Задръжте позицията за секунда или две
Върнете се в горната част на движението, докато стискате подколенните сухожилия
Работата с тези мускули ще помогне за улесняване на движенията при клякане, скокове или огъване на гръбначния стълб.
Отказ от отговор на публикацията в д-р Алекс Хименес
Обхватът на нашата информация е ограничен до хиропрактика, мускулно-скелетна система, физически лекарства, уелнес и чувствителни здравни проблеми и / или статии, теми и дискусии за функционална медицина. Използваме функционални протоколи за здраве и уелнес за лечение и подпомагане на грижи при наранявания или разстройства на опорно-двигателния апарат. Нашите публикации, теми, теми и прозрения обхващат клинични въпроси, проблеми и теми, които се отнасят и подпомагат пряко или косвено нашата клинична практика. *
Нашият офис направи разумен опит да предостави подкрепящи цитати и идентифицира съответното изследователско проучване или изследвания, подкрепящи нашите публикации. Ние също така правим копия на подкрепящи изследователски изследвания, достъпни за борда и / или обществеността при поискване. Разбираме, че обхващаме въпроси, които изискват допълнително обяснение как може да подпомогне конкретен план за грижи или протокол за лечение; следователно, за да обсъдите допълнително темата по-горе, моля не се колебайте да попитате д-р Алекс Хименес или да се свържете с нас на 915-850-0900. Доставчикът (ите) Лицензиран (и) в Тексас и Ню Мексико *
Храните, които ядем, могат да имат потенциал да бъдат полезни или вредни за здравето ни. Лошото хранене може да причини различни здравословни проблеми, включително затлъстяване, сърдечно-съдови заболявания и диабет тип 2. Междувременно правилното хранене може да ви накара да се почувствате заредени с енергия, да намали риска от здравословни проблеми, както и да ви помогне да поддържате и регулирате здравословното тегло. Ако искате да насърчите дълголетието, трябва да подхранвате тялото си с добри храни. В следващата статия ще изброим няколко добри храни, които в крайна сметка могат да помогнат за насърчаване на дълголетието, като също така помагат за подобряване на цялостното здраве и благосъстояние.
Кръстосано зеленчуци
Кръстоцветните зеленчуци имат уникалната способност да променят хормоните ни, да задействат естествената система за детоксикация на организма и дори да намалят растежа на раковите клетки. Те трябва да се дъвчат старателно или да се ядат настъргани, нарязани, изцедени или смесени, за да се освободят полезните им свойства. Установено е също, че сулфорафанът, намиращ се в кръстоцветните зеленчуци, помага да се предпази стената на кръвоносните съдове от възпаление, което може да причини сърдечни заболявания. Кръстоцветните зеленчуци като къдраво зеле, зеле, брюкселско зеле, карфиол и броколи са едни от най-богатите на хранителни вещества храни в света.
Зелена салата
Суровите листни зелени имат по-малко от 100 калории на килограм, което ги прави идеалната храна за отслабване. Яденето на повече зелени салати се свързва и с намаления риск от сърдечен удар, инсулт, диабет и няколко вида ракови заболявания. Суровите листни зеленини също са богати на основния В-витамин фолат, плюс лутеин и зеаксантин, каротеноиди, които могат да помогнат за защита на очите. Мастноразтворимите фитохимикали, като каротеноиди, които се съдържат в зелените салати като маруля, спанак, къдраво зеле, горчица и горчица, също имат антиоксидантни и противовъзпалителни ефекти в организма.
ядки
Ядките са нискогликемична храна и чудесен източник на здравословни мазнини, растителни протеини, фибри, антиоксиданти, фитостероли и минерали, което също помага да се намали гликемичното натоварване на цялото хранене, което ги прави съществена част от антидиабета диета. Независимо от тяхната калорична плътност, яденето на ядки може да помогне за насърчаване на загубата на тегло. Ядките също могат да намалят холестерола и да помогнат за намаляване на риска от сърдечни заболявания.
Семена
Семената, подобно на ядките, също осигуряват здравословни мазнини, антиоксиданти и минерали, но те имат повече протеин и са богати на микроелементи. Чиа, лен и конопено семе са богати на омега-3 мазнини. Чиа, лен и сусамови семена също са богати лигнани или фитоестрогени, борещи се с рак на гърдата. Освен това, сусамовите семена са богати на калций и витамин Е, а тиквените семки са богати на цинк.
Зърната
Плодовете са богати на антиоксиданти плодове, които могат да помогнат за укрепване на здравето на сърцето. Изследователски проучвания, при които участниците ядат ягоди или боровинки ежедневно в продължение на няколко седмици, отчитат подобрения в кръвното налягане, общия и LDL холестерола и дори признаци на оксидативен стрес. Плодовете имат също и противоракови свойства и е доказано, че помагат за предотвратяване на когнитивен спад, свързан със стареенето.
Нар
Най-известният фитохимикал в наровете, пуникалагин, е отговорен за повече от половината от антиоксидантната активност на плода. Фитохимикалите от нар имат противоракови, кардиопротективни и полезни за мозъка ползи. В едно изследователско проучване по-възрастните хора, които пият сок от нар ежедневно в продължение на 28 дни, се представят по-добре при тест за памет в сравнение с тези, които пият плацебо напитка.
Боб
Яденето на боб и други бобови растения може да помогне за балансиране на кръвната захар, да намали апетита ви и да предпази от рак на дебелото черво. Фасулът е храна против диабет, която може да помогне за насърчаване на загубата на тегло, защото се усвоява бавно, което забавя повишаването на кръвната захар след хранене и спомага за предотвратяване на желанието за храна, като насърчава ситостта. Яденето на боб и други бобови растения два пъти седмично е установено, че намалява риска от рак на дебелото черво. Яденето на боб и други бобови растения, като червен боб, черен боб, нахут, леща и цепена грах, също осигурява значителна защита срещу други видове рак.
Гъби
Яденето на гъби редовно е свързано с намален риск от рак на гърдата. Белите гъби и Портобело са особено полезни срещу рака на гърдата, тъй като имат инхибитори на ароматазата или съединения, които инхибират производството на естроген. Гъбите са показали, че имат противовъзпалителни ефекти, както и осигуряват засилена активност на имунните клетки, предотвратяване на увреждането на ДНК, забавяне на растежа на раковите клетки и инхибиране на ангиогенезата. Гъбите винаги трябва да се готвят, тъй като суровите гъби имат потенциално канцерогенен химикал, известен като агаритин, който се намалява значително при готвене.
Лук и чесън
Лукът и чесънът осигуряват ползи за сърдечносъдовата и имунната система, както и антидиабетни и противоракови ефекти. Те също са свързани с по-малък риск от рак на стомаха и простатата. Лукът и чесънът са известни със своите органични съединения, които помагат да се предотврати развитието на рак чрез детоксикация на канцерогени, намаляване на растежа на раковите клетки и блокиране на ангиогенезата. Лукът и чесънът също имат високи концентрации на стимулиращи здравето флавоноидни антиоксиданти, които имат противовъзпалителни ефекти, които могат да помогнат за осигуряване на превенция на рака.
домати
Доматите са богати на различни хранителни вещества, като ликопен, витамин С и Е, бета-каротин и флавонолни антиоксиданти. Ликопенът може да помогне за предпазване от рак на простатата, UV увреждане на кожата и? сърдечно-съдови заболявания. Ликопенът се усвоява по-добре, когато се готвят домати. Една чаша доматен сос съдържа около 10 пъти количеството ликопен като чаша сурови, нарязани домати. Също така имайте предвид, че каротеноидите, като ликопен, се усвояват най-добре, когато са придружени от здравословни мазнини, затова се наслаждавайте на доматите си в салата с ядки или дресинг на ядкова основа за допълнителни хранителни ползи.
Храните, които ядем, могат да имат потенциал да бъдат полезни или вредни за нашето здраве. Лошото хранене може да причини различни здравословни проблеми, включително затлъстяване, сърдечно-съдови заболявания и диабет тип 2. Междувременно правилното хранене може да ви накара да се чувствате енергични, да намали риска от здравословни проблеми, както и да ви помогне да поддържате и регулирате здравословно тегло. Ако искате да насърчите дълголетието, трябва да подхранвате тялото си с добри храни. Добрите храни също могат да помогнат за намаляване на възпалението, свързано с различни здравословни проблеми, включително болки в ставите и артрит. Здравните специалисти, като хиропрактики, могат да предложат съвети за диета и начин на живот, за да помогнат за насърчаване на здравето и благосъстоянието. В следващата статия ще изброим няколко добри храни, които в крайна сметка могат да помогнат за насърчаване на дълголетието. – Д-р Алекс Хименес DC, CCST Insight
Zesty сок от цвекло
Порции: 1 Време за готвене: 5-10 минути
1 грейпфрут, обелен и нарязан
1 ябълка, измита и нарязана на филийки
1 цяло цвекло и листа, ако ги имате, измити и нарязани на филийки
1-инчово копче джинджифил, изплакнато, обелено и нарязано
Изцедете всички съставки в висококачествена сокоизстисквачка. Най-добре се сервира веднага.
Само един морков ви дава целия дневен прием на витамин А
Да, консумацията на само един варен морков от 80 g (2 унции) ви дава достатъчно бета каротин, за да може тялото ви да произведе 1,480 микрограма (mcg) витамин А (необходим за обновяване на клетките на кожата). Това е повече от препоръчителния дневен прием на витамин А в Съединените щати, който е около 900 mcg. Най-добре е да ядете моркови варени, тъй като това омекотява клетъчните стени, позволявайки на повече бета каротин да се абсорбира. Добавянето на по-здравословни храни към вашата диета е чудесен начин да подобрите цялостното си здраве.
Обхватът на нашата информация е ограничен до хиропрактика, мускулно-скелетна система, физически лекарства, уелнес и чувствителни здравни проблеми и / или статии, теми и дискусии за функционална медицина. Използваме функционални протоколи за здраве и уелнес за лечение и подпомагане на грижи при наранявания или разстройства на опорно-двигателния апарат. Нашите публикации, теми, теми и прозрения обхващат клинични въпроси, проблеми и теми, които се отнасят и подкрепят пряко или косвено нашия клиничен обхват на практика. * Нашият офис направи разумен опит да предостави подкрепящи цитати и идентифицира съответното изследване или проучвания в подкрепа на нашите публикации. Ние също така правим копия на подкрепящи изследователски изследвания, достъпни за борда и / или обществеността при поискване. Разбираме, че обхващаме въпроси, които изискват допълнително обяснение за това как може да помогне в определен план за грижи или протокол за лечение; следователно, за да обсъдите допълнително темата по-горе, моля не се колебайте да попитате д-р Алекс Хименес или да се свържете с нас на 915-850-0900. Доставчикът (ите) Лицензиран (и) в Тексас * и Ню Мексико *
Куриран от д-р Алекс Хименес DC, CCST
Литература:
Джоел Фурман, д-р. 10 най-добри храни, които можете да ядете, за да живеете по-дълго и да останете здрави Healthwell Health, 6 юни 2020 г., www.verywellhealth.com/best-foods-for-longevity-4005852.
Даудън, Анджела. „Кафето е плод и други невероятно истински хранителни факти“ MSN Lifestyle, 4 юни 2020 г., www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
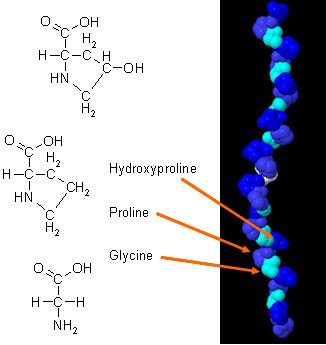
Ако изпитвате някоя от тези ситуации, тогава колагеновите ви пептиди може да са ниски.
Има бяха нови проучвания за това как колагенът може да подобри състава на тялото, когато се комбинира с ежедневни упражнения. Колагенът в тялото има уникален аминокиселинен състав, който играе съществена роля в анатомията на тялото. Колагенният протеин е концентриран източник на глицин, пролин и хидроксипролин и когато се сравнява с всички други диетични протеини, той прави колагена потенциален практичен избор като структурен протеин.
In проучване 2015, изследователите са показали как ефикасните добавки с колаген могат да подобрят телесния състав при активни мъже. Резултатите показват как всеки мъж участва в тренировки с тежести поне три пъти седмично и трябва да добавя поне 15 грама колагенови пептиди, за да постигне максимално здраве. Оценките, които тестът предоставя, са тест за сила, анализ на биоимпеданс (BIA) и мускулни биопсии. Тези тестове гарантират, че мъжете се представят добре след приемане на добавките с колаген, а резултатите показват как телесната им маса е имала увеличение на телесна маса без мазнини. Друго проучване показа как добавянето на колаген протеин, когато се комбинира с тренировки за съпротива, които могат да повишат мускулната маса и мускулната сила при възрастни хора, както и при хора със саркопения.
Полезни свойства с колаген
Има много полезни свойства които добавките с колаген могат да осигурят на тялото, когато се консумират. Има хидролизиран колаген и желатин и могат да помогнат за подобряване на структурата на кожата на човек. Въпреки че няма много проучвания за добавките с колаген, има отлични обещания за зоните по тялото. Те са:
Мускулна маса: Добавките с колаген, когато се комбинират със силови тренировки, могат да увеличат мускулната маса и силата в тялото.
Артрит: Добавките с колаген могат да помогнат на хора с остеоартрит. Проучванията показват че когато хората с остеоартрит приемат добавки с колаген, те откриват огромен спад в болката, която изпитват.
Еластичност на кожатаВ проучване 2014, в него се посочва, че жените, които са приемали добавки с колаген и са показали подобрения в еластичността на кожата. Колагенът може да се използва и при локални лечения, за да помогне за подобряване на външния вид на кожата на човек чрез минимизиране на фините линии и бръчки.
Не само добавките с колаген осигуряват полезни свойства на специфичните области на тялото, но има четири основни типа колаген и каква е тяхната роля в човешкото тяло, както и функциите им:
Въведете 1: Колагенът тип 1 отчита 90% от колагена на тялото и се състои от плътно опаковани влакна, които осигуряват структури на кожата, костите, съединителната тъкан и зъбите, които са в тялото.
Въведете 2: Колагенът тип 2 е изграден от свободно опаковани влакна, които се намират в еластичния хрущял, който помага за омекотяване на ставите в тялото.
Въведете 3: Колагенът тип 3 помага за поддържането на структурата на мускулите, органите и артериите, които гарантират, че тялото функционира правилно.
Въведете 4: Колагенът тип 4 се намира в слоевете на кожата на всеки и помага за филтрирането в тялото.
Тъй като тези четири вида колаген се намират в тялото, важно е да се знае, че колагенът може естествено да намалява с времето с възрастта, тъй като тялото ще произвежда по-ниско качество на колагена. Един от видимите признаци на намаляване на колагена е, когато кожата на човешкото тяло става по-малко твърда и еластична, както и отслабването на хрущяла поради стареенето.
Фактори, които могат да увредят колагена
Въпреки че колагенът може да намалява естествено с възрастта, много фактори могат да унищожат колагените, които са вредни за кожата. Вредните фактори могат да включват:
Захар и въглехидрати: Рафинирани захари и въглехидрати може да пречи със способността на колагена да се самовъзстановява върху кожата. Така че чрез минимизиране на консумацията на захар и въглехидрати в тялото, той може да намали ефектите от съдова, бъбречна и кожна тъканна дисфункция.
Излагане на слънце: Въпреки че получаването на достатъчно слънце може да помогне на човек да се наслади на деня, излагането на слънце за продължителен период от време може да причини повреда към кожата и унищожават колагеновите пептиди. Ефектите от прекомерното излагане на слънце могат да причинят фотостареене на кожата и да предизвикат оксидативен стрес в тялото.
пушене: Когато човек пуши, може намаляване на производството на колаген в тялото, което кара тялото да има преждевременни бръчки и ако тялото е наранено, лечебният процес ще бъде по-бавен и може да доведе до заболявания в тялото.
автоимунни заболявания: Някои автоимунни заболявания също могат да увредят производството на колаген като лупус.
Заключение
Колагенът е жизненоважен за тялото, тъй като помага на кожата да бъде нежна и стегната. Естествено, той ще намалее с напредването на възрастта, така че приемането на колагенови добавки може да гарантира, че тялото може да функционира правилно. Когато вредните фактори влияят върху тялото, те могат да спрат или дори да увредят производството на колаген и да ускорят процеса на образуване на преждевременни бръчки, което кара човек да изглежда по-възрастен, отколкото е. някои продукти може да подпомогне клетъчната активност на тялото, като осигури по-добра стабилност, бионаличност и храносмилателен комфорт.
Обхватът на нашата информация е ограничен до хиропрактика, проблеми с мускулно-скелетното и нервно здраве или статии, теми и дискусии за функционалната медицина. Използваме функционални здравни протоколи за лечение на наранявания или нарушения на опорно-двигателния апарат. Нашият офис направи разумен опит да предостави подкрепящи цитати и определи съответното изследователско проучване или проучвания в подкрепа на нашите публикации. Ние също така правим копия на подкрепящи изследователски проучвания на разположение на съвета и на обществеността при поискване. За да обсъдим допълнително темата по-горе, моля, попитайте д-р Алекс Хименес или се свържете с нас на 915-850-0900.
Литература:
Бош, Рикардо и др. Механизми на фотостареене и кожна фотокарциногенеза и фотозащитни стратегии с фитохимикали. Антиоксиданти (Базел, Швейцария), MDPI, 26 март 2015 г., www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Данби, Ф. Уилям. Хранене и стареене на кожата: захар и гликация. Клиники по дерматология, Национална медицинска библиотека на САЩ, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Дженингс, Кери-Ан. Колаген – какво е това и за какво е полезен? Healthline, 9 септември 2016 г., www.healthline.com/nutrition/collagen.
Юргелевич, Майкъл. Ново проучване демонстрира ползите от колагеновите пептиди за подобряване на състава на тялото, съчетано с упражнения. Проекти за здраве, 31 май 2019 г., blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. Пушенето засяга синтеза на колаген и оборота на извънклетъчния матрикс в човешката кожа. Британското списание по дерматология, Национална медицинска библиотека на САЩ, апр. 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, et al. Пероралната добавка на специфични колагенови пептиди има благоприятен ефект върху физиологията на човешката кожа: двойно-сляпо, плацебо-контролирано проучване. Кожна фармакология и физиология, Национална медицинска библиотека на САЩ, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, et al. Ефект на новия хидролизиран екстракт от пилешки гръден хрущял с ниско молекулно тегло, BioCell Collagen, върху подобряване на симптомите, свързани с остеоартрит: рандомизирано, двойно-сляпо, плацебо-контролирано проучване. Вестник на селскостопанската и хранителната химия, Национална медицинска библиотека на САЩ, 25 Apr. 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise и др. Добавката с колагенови пептиди в комбинация с тренировки за съпротива подобрява състава на тялото и повишава мускулната сила при възрастни мъже със саркопея: рандомизирано контролирано проучване. Британският вестник на храненето, Cambridge University Press, 28 октомври 2015 г., www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Като информира хората за това как Националният университет по здравни науки предоставя знания за бъдещите поколения, университетът предлага голямо разнообразие от медицински професии за функционална медицина.
Все едно сте били диагностицирани с цьолиакия, синдром на раздразнените черва, дивертикулоза/дивертикулит или синдром на пропускливи черва?
Прекомерно оригване, оригване или подуване на корема?
Ненормално подуване след определени пробиотици или естествени добавки?
Подозрение за хранителна малабсорбция?
Отслабват ли храносмилателните проблеми с отпускането?
Ако изпитвате някоя от тези ситуации, тогава може да имате проблеми с червата и може да се наложи да опитате програмата 4R.
Хранителната чувствителност, ревматоидният артрит и тревожността се свързват с нарушена стомашно-чревна пропускливост. Тези различни състояния могат да възникнат от много фактори, които могат да повлияят на храносмилателния тракт. Ако не се лекува, това потенциално може да бъде резултат от дисфункция на чревната пропусклива бариера, причинявайки възпаление и тежки здравословни състояния, които могат да развият червата. Програмата 4R се използва за възстановяване на здрави черва в тялото и включва четири стъпки. Те са: премахване, подмяна, реинокулация и ремонт.
Чревна пропускливост
Чревната пропускливост помага за защита на тялото и гарантира, че вредните бактерии не навлизат в червата. Предпазва тялото от потенциални фактори на околната среда които могат да бъдат вредни и навлизат през храносмилателния тракт. Това може да бъде или токсин, патогенни микроорганизми и други антигени, които могат да навредят на храносмилателния тракт, причинявайки проблеми. Лигавицата на червата се състои от слой от епителни клетки, които са разделени с тесни връзки. В здрави черва, плътната връзка регулира чревната пропускливост, като селективно позволява на веществата да влизат и преминават през чревната бариера и предотвратява абсорбирането на вредни фактори.
Определени фактори на околната среда могат да повредят плътната връзка и резултатът е, че може да увеличи чревната пропускливост, което причинява чревна хиперпропускливост или пропускливост на червата в тялото. Допринасящите фактори могат да увеличат чревната пропускливост като прекомерно количество наситени мазнини и алкохол, дефицит на хранителни вещества, хроничен стрес и инфекциозни заболявания.
С повишена чревна пропускливост в червата, той може да даде възможност на антигените да преминат през чревната лигавица и да влязат в кръвния поток, причинявайки имунен отговор и възпаление на тялото. Има определени стомашно-чревни състояния, които са свързани с чревна хиперпропускливост и ако не се лекуват, могат да предизвикат определени автоимунни състояния, които могат да причинят вреда на тялото.
Програма 4Rs
4Rs е програма, която здравните специалисти съветват пациентите си да използват, когато се справят с разрушителни храносмилателни проблеми и подпомагат заздравяването на червата.
Премахване на проблема
Първата стъпка в програмата 4Rs е премахването на вредни патогени и провокатори на възпаление, които са свързани с повишена чревна пропускливост. Тригери като стрес и хронична консумация на алкохол могат да навредят много на тялото на индивида. Така че насочването към тези вредни фактори от тялото е да се лекува с лекарства, антибиотици, добавки и се препоръчва премахването на възпалителни храни от диетата, включително:
- Алкохол
- Глутен
- Хранителни добавки
– Нишестета
– Някои мастни киселини
– Някои храни, към които човек е чувствителен
Замяна на хранителните вещества
Втората стъпка от програмата 4Rs е замяната на хранителните вещества, които причиняват проблеми с червата чрез възпаление. Някои хранителни вещества могат да помогнат за намаляване на възпалението в червата, като същевременно гарантират, че храносмилателният тракт се поддържа. Има някои противовъзпалителни храни, които са питателни. Те включват:
– Храни с високо съдържание на фибри
– Омега-3
- Зехтин
– гъби
– противовъзпалителни билки
Има някои добавки, които могат да се използват за подпомагане на храносмилателната функция, като подпомагат и усвояват хранителните вещества за насърчаване на здрави черва. Това, което правят храносмилателните ензими, е, че подпомагат разграждането на мазнините, протеините и въглехидратите в червата. Това ще помогне на хора, които имат увреден храносмилателен тракт, непоносимост към храни или страдащи от цьолиакия. Добавки като добавки с жлъчна киселина могат да помогнат за усвояването на хранителните вещества чрез сливане на липидите заедно. Изследванията са заявили че жлъчните киселини са били използвани за лечение на черния дроб, жлъчния мехур и жлъчните пътища, като същевременно предотвратяват образуването на камъни в жлъчката след бариатрична операция.
Реинокулира червата
Третата стъпка е от програмата 4rs за реинокулиране на чревния микроб с полезни бактерии за насърчаване на здравословната функция на червата. Показани са изследвания че пробиотичните добавки са били използвани за подобряване на червата чрез възстановяване на полезните бактерии. С тези добавки те осигуряват подобрение на червата чрез отделяне на противовъзпалителни вещества в тялото, подпомагат поддържането на имунната система, променят микробния състав на тялото и намаляват чревната пропускливост в чревната система.
Тъй като намират се пробиотици във ферментирали храни и се считат за преходни, тъй като не са устойчиви в стомашно-чревния тракт и са полезни. Изненадващо, те все още оказват влияние върху човешкото здраве поради влияние върху червата, като произвеждат витамини и антимикробни съединения, като по този начин осигуряват разнообразие и функция на червата.
Възстановяване на червата
Последната стъпка от програмата 4Rs е ремонт на червата. Тази стъпка включва възстановяване на чревната лигавица на червата със специфични хранителни вещества и билки. Тези билки и добавки могат да помогнат за намаляване на чревната пропускливост и възпаление в тялото. Някои от тези билки и добавки включват:
- Алое вера
– Хиос мастична дъвка
– DGL (деглициризиран женско биле)
– Корен от бяла ружа
– L-глутамин
– Омега-3
Полифеноли
– витамин D
- Цинк
Заключение
Тъй като много фактори могат да повлияят неблагоприятно на храносмилателната система по вреден начин и могат да допринесат за няколко здравословни състояния. Основната цел на програмата 4Rs е да минимизира тези фактори, които увреждат червата и намаляват възпалението и повишената чревна пропускливост. Когато пациентът се запознае с полезните фактори, които предоставят 4Rs, това може да доведе до здрави, излекувани черва. някои продукти са тук, за да подпомогнат стомашно-чревната система, като поддържат червата, подобряват метаболизма на захарта и се насочват към аминокиселините, които са предназначени да поддържат червата.
Обхватът на нашата информация е ограничен до хиропрактика, проблеми с мускулно-скелетното и нервно здраве или статии, теми и дискусии за функционалната медицина. Използваме функционални здравни протоколи за лечение на наранявания или нарушения на опорно-двигателния апарат. Нашият офис направи разумен опит да предостави подкрепящи цитати и определи съответното изследователско проучване или проучвания в подкрепа на нашите публикации. Ние също така правим копия на подкрепящи изследователски проучвания на разположение на съвета и на обществеността при поискване. За да обсъдим допълнително темата по-горе, моля, попитайте д-р Алекс Хименес или се свържете с нас на 915-850-0900.
Литература:
Де Сантис, Стефания и др. Хранителни ключове за модулация на чревната бариера. Граници в имунологията, Frontiers Media SA, 7 декември. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Яниро, Джанлука и др. Добавяне на храносмилателни ензими при стомашно-чревни заболявания. Актуален метаболизъм на лекарствата, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui и др. „Пропускане на червата като сигнал за опасност за автоимунни заболявания Frontiers, Frontiers, 5 май 2017 г., www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon и др. Ферментиралите храни като хранителен източник на живи организми. Граници в микробиологията, Frontiers Media SA, 24 август 2018 г., www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., et al. Бързото нарушаване на функцията на чревната бариера от глиадин включва променена експресия на апикални свързващи протеини. FEBS Натиснете, John Wiley & Sons, Ltd, 8 август 2005 г., febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Сартор, Р Балфур. Терапевтична манипулация на чревната микрофлора при възпалителни чревни заболявания: антибиотици, пробиотици и пребиотици. Gastroenterology, Национална медицинска библиотека на САЩ, май 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Хроничната болка е често срещан здравословен проблем, който засяга много хора в Съединените щати. Докато няколко медицински състояния, като фибромиалгия и синдром на миофасциална болка, могат да причинят хронична болка, тя може да се развие и поради различни други здравословни проблеми. Изследванията показват, че широко разпространеното възпаление е водещата причина за хронична болка. Възпалението е естествен защитен механизъм срещу нараняване, заболяване или инфекция. Но ако възпалителният процес продължи твърде дълго, той може да стане проблематичен.
Възпалението сигнализира имунната система да лекува и възстановява увредената тъкан, както и да се предпазва от бактерии и вируси. Както е споменато по-горе, обаче, хроничното възпаление може да предизвика различни здравни проблеми, включително симптоми на хронична болка. Модификациите на здравословния начин на живот могат да помогнат при управлението на хроничната болка, но първо трябва да разберем общите причини за хроничната болка.
Какво е остро възпаление?
Острото възпаление, например, възниква след нараняване или нещо толкова просто като възпалено гърло. Това е естествен отговор с неблагоприятни ефекти, което означава, че работи локално в региона, където е намерен здравният проблем. Общите признаци на остро възпаление включват подуване, зачервяване, топлина, болка и загуба на функция, както е посочено от Националната библиотека по медицина. Когато се развие остро възпаление, кръвоносните съдове се разширяват, което води до увеличаване на притока на кръв, а белите кръвни клетки в увредената област подпомагат възстановяването.
По време на тежко възпаление, съединенията, наречени цитокини, се освобождават от увредената тъкан. Цитокините действат като "аварийни сигнали", които довеждат до собствените имунни клетки на човешкото тяло, както и до хормони и множество хранителни вещества, за да се поправи здравния проблем. Освен това, хормон-подобни вещества, известни като простагландини, причиняват образуването на кръвни съсиреци, за да лекуват повредената тъкан, и те също могат да предизвикат треска и болка като част от възпалителната процедура. Тъй като увреждането или нараняване се възстановява, възпалението намалява.
Какво е хронично възпаление?
За разлика от острото възпаление, хроничното възпаление има дълготрайни ефекти. Хроничното възпаление, известно също като персистиращо възпаление, произвежда ниски нива на възпаление в човешкото тяло, както се вижда от увеличаването на маркерите на имунната система, разположени в кръвта и клетъчните тъкани. Хроничното възпаление може също да предизвика прогресиране на различни заболявания и състояния. Повишените нива на възпаление могат понякога да задействат дори ако няма нараняване, заболяване или инфекция, които също могат да причинят реакция на имунната система.
В резултат на това имунната система на човешкото тяло може да започне да атакува здрави клетки, тъкани или органи. Изследователите все още се опитват да разберат последствията от хроничното възпаление в човешкото тяло и механизмите, включени в този естествен защитен процес. Например, хроничното възпаление е свързано с различни здравословни проблеми, като сърдечни заболявания и инсулт.
Една теория предполага, че когато възпалението остане в кръвоносните съдове, то може да насърчи натрупването на плака. Според Американската асоциация по сърдечни заболявания или AHA, ако имунната система идентифицира плаката като чуждестранен нашественик, белите кръвни клетки могат да се опитат да затворят плаката, намерена в кръвта, преминаваща през артериите. Това може да създаде кръвен съсирек, който може да блокира притока на кръв към сърцето или мозъка, което води до неговата нестабилност и разкъсване. Ракът е друг здравен проблем, свързан с хронично възпаление. Освен това, според Националния институт за рака, увреждането на ДНК може да бъде причинено и от хронично възпаление.
Устойчивото, нискокачествено възпаление често няма никакви симптоми, но здравните специалисти могат да проверят за С-реактивен протеин или CRP, известен като липоева киселина, маркер за възпаление, открит в кръвта. Повишените нива на CRP са свързани с повишен риск от сърдечносъдови заболявания. Повишени нива на CRP могат да бъдат открити при хронични заболявания като лупус или ревматоиден артрит.
В случай на други хронични състояния, като фибромиалгия, нервната система реагира на специфична стимулация, но това е възпаление, което причинява симптоми на хронична болка. Субективно е почти невъзможно да се каже разликата между хроничната болка, причинена от свръхчувствителната нервна система и хроничната болка, причинена от широко разпространено възпаление. Освен търсенето на улики в кръвния поток, храненето на човека, навиците на начина на живот и експозицията на околната среда също могат да насърчат хроничното възпаление.
Възпалението е естественият защитен механизъм на имунната система срещу нараняване, заболяване или инфекция. Докато този възпалителен отговор може да помогне за излекуване и възстановяване на тъканите, хроничното, широко разпространено възпаление може да причини различни здравословни проблеми, включително симптоми на хронична болка. Балансиран храненето, включително разнообразни диети и гладуване, може да помогне за намаляване на възпалението. Гладуването, известно още като ограничаване на калориите, насърчава апоптозата на клетките и възстановяването на митохондриите. Диетата, имитираща гладуването, която е част от диетата за дълголетие, е диетична програма, която „подвежда” човешкото тяло в състояние на гладно, за да изпита предимствата на традиционното гладуване. Преди да следвате някоя от диетите, описани в тази статия, не забравяйте да се консултирате с лекар.
Д-р Алекс Хименес DC, CCST Insight
Хранене, диети, пост и хронична болка
Противовъзпалителните диети се състоят основно от консумация на пресни плодове и зеленчуци, риба и мазнини. Средиземноморската диета е например противовъзпалителна диета, която насърчава консумацията на умерени количества ядки, поглъщане на много малко месо и пиене на вино. Противовъзпалителните хранителни части, като омега-3 мастни киселини, предпазват човешкото тяло от daмагьосник предизвикано от възпаление.
Противовъзпалителната диета включва също така да се пази от храни, които биха могли да стимулират възпалението. Той е идеален за намаляване на количеството храни, които консумирате с високо съдържание на транс и наситени мазнини, като например месо. Освен това, противовъзпалителната диета ограничава консумацията на рафинирани въглехидрати и храни, като хляб и ориз. Те също така насърчават намаляването на използването на маргарин и масла, които са опаковани с омега-6 мастни киселини, като слънчоглед, шафран намлява царевични масла.
Известно е, че гладуването или ограничаването на калориите намалява оксидативния стрес и забавя механизмите на стареене в различни организми. Ефектите от гладуването включват програмирана клетъчна смърт или апоптоза, транскрипция, мобилна енергийна ефективност, митохондриална биогенеза, антиоксидантни механизми и циркаден ритъм. Постенето също допринася за митохондриалната аутофагия, известна като митофагия, където гените в митохондриите се стимулират да се подлагат на апоптоза, която насърчава възстановяването на митохондриите.
Периодичното гладуване може да ви помогне да се борите с възпалението, да подобрите храносмилането и да увеличите дълголетието си. Човешкото тяло е проектирано така, че да може да оцелява продължително време без храна. Изследвания показват, че периодичното гладуване може да има положителни промени в цялостния състав на чревната ви микробиота. Освен това, периодичното гладуване може да намали инсулиновата резистентност, като същевременно увеличава отговора на имунната система. И накрая, периодичното гладуване може да насърчи производството на вещество, известно като? -Хидроксибутират, което блокира част от имунната система, участваща в възпалителни заболявания, както и значително намаляване на производството на възпалителни маркери, като цитокини и С-реактивен протеин или CRP, споменати по-горе.
Планът за хранене за дълголетие, представен в книгата на д-р Валтер Лонго, елиминира консумацията на преработени храни, които могат да причинят възпаление, насърчавайки благосъстоянието и дълголетието. Тази уникална диетична програма, за разлика от повечето традиционни диети, не насърчава загуба на тегло. Въпреки че може да имате намаление на теглото, акцентът на тази уникална диетична програма е върху здравословното хранене. Диетичният план за дълголетие е доказал, че помага да се активира обновяването на стволовите клетки, да се намали абдоминалната мазнина и да се предотврати загуба на кости и мускули, свързани с възрастта, както и да се повиши устойчивостта към развитие на сърдечно-съдови заболявания, болест на Алцхаймер, диабет и рак.
Диета, която имитира гладно, или FMD, ви позволява да изпитате ползите от традиционното гладно, без да лишавате тялото си от храна. Основната разлика на FMD е, че вместо да елиминира напълно цялата храна за няколко дни или дори седмици, вие ограничавате приема на калории само за пет дни от месеца. Шап може да се практикува веднъж месечно, за да се подпомогне цялостното здраве и добро здраве.
Докато всеки може да проследи FMD самостоятелно ProLon Диета, която имитира гладно, предлага програма за хранене 5-ден, която е индивидуално опакована и етикетирана за всеки ден, която обслужва храните, от които се нуждаете за шап, в точни количества и комбинации. Програмата за хранене се състои от готови за консумация или лесни за приготвяне растителни храни, включително барове, супи, леки закуски, добавки, концентрат за напитки и чайове. Преди да започнете Пролонова диета, имитираща гладно, 5-дневна програма за храненеили някой от промените в начина на живот, описани по-горе, моля, уверете се, че говорете със здравен специалист, за да разберете коя терапия за хронична болка е подходяща за вас.
Обхватът на нашата информация е ограничен до статии, теми и дискусии по хиропрактика, проблеми със здравето на гръбначния стълб и функционална медицина. За допълнително обсъждане на темата по-горе, моля не се колебайте да попитате д-р Алекс Хименес или да се свържете с нас на адрес 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителна дискусия по темата: Остра болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за увреждане и пропуснати дни по време на работа по целия свят. Болката в гърба се свързва с втората най-често срещана причина за посещения на лекарски кабинет, превъзхождана само от горните дихателни инфекции. Приблизително 80 процента от населението ще изпита болки в гърба поне веднъж през целия си живот. Вашият гръбначен стълб е сложна структура, съставена от кости, стави, сухожилия и мускули сред другите меки тъкани. Наранявания и / или утежнени състояния, като например херния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
КСИМОГЕНИ Ексклузивни професионални формули са достъпни чрез избрани лицензирани професионалисти в областта на здравеопазването. Продажбата по интернет и дисконтирането на формулите XYMOGEN са строго забранени.
Гордост, Д-р Александър Хименес прави формулите XYMOGEN достъпни само за пациенти под нашата грижа.
Моля, обадете се в нашия офис, за да можем да назначим консултация с лекар за незабавен достъп.
Ако сте пациент Медицинска и хиропрактична клиника за нараняване, можете да попитате за XYMOGEN, като се обадите 915-850-0900.
За ваше удобство и преглед на XYMOGEN продукти, моля прегледайте следната връзка. *XYMOGEN-подреждане иИзтегли
* Всички горепосочени политики на XYMOGEN остават в сила.
Придържането към специфична диета за поддържане на правилното хранене понякога може да направи храненето стресиращо. Естествените промени в начина на живот са ключът към промяната на хранителните навици и това може да ви помогне да живеете по-дълъг и по-здравословен живот. Планът за хранене за дълголетие, създаден от д-р Валтер Лонго, е серия от практически насоки за хранене, които се фокусират върху промяната на хранителните ви начини за постигане на цялостно здраве и уелнес.
Правилата на плана за хранене за дълголетие
Като следвате хранителните съвети по-долу, можете да преразгледате сегашния си план за хранене и да започнете да ядете по-здравословно, без да се налага стрес от традиционната диета. Планът за хранене за дълголетие елиминира консумацията на преработени храни, които могат да предизвикат различни здравни проблеми и увеличава консумацията на хранителни вещества, които насърчават дълголетието. Тази уникална диетична програма споделя резултатите от приблизително 25 години на научни изследвания на едно просто решение, което може да помогне на хората да изпитат общото благосъстояние чрез правилното хранене.
Въпреки това, за разлика от повечето традиционни диети, Планът за диета за дълголетие не насърчава загубата на тегло. Въпреки че може да изпитате намаляване на теглото, акцентът на тази уникална диетична програма е върху по-здравословното хранене. Диетичният план за дълголетие е демонстриран, за да ви помогне да активирате обновяването на основата на стволови клетки, да отслабнете и да намалите мазнините в корема, да предотвратите загуба на кост и мускули, свързана с възрастта, да изградите устойчивост към развитие на сърдечно-съдови заболявания, болест на Алцхаймер, диабет и рак, както и като удължават дълголетието. По-долу ще обобщим 8-те най-често срещани хранителни съвета от Диетата за дълголетие, които в крайна сметка могат да помогнат за по-дълъг и по-здравословен живот.
Диетичният план за дълголетие е уникална диетична програма, разработена от д-р Валтер Лонго за насърчаване на цялостното здраве, уелнес и дълголетие. Чрез прости промени в начина на живот, хората могат да променят хранителните си навици и да се възползват от многото ползи за здравето на тази диетична програма. Следвайки пескатарска диета и следвайки Пролонова диета, имитираща гладуванеСред другите хранителни съвети, описани по-долу, хората могат да живеят по-дълго и по-здравословно. Традиционните диети често могат да бъдат трудни и стресиращи, но планът за диета за дълголетие е практична и уникална диетична програма, която може да бъде подходяща за много хора.
Д-р Алекс Хименес DC, CCST Insight
Хранителни съвети 8 на плана за диета за дълголетие
Следвайте диета на Pescatarian
Като част от плана за хранене на дълголетието, следвайте диета на пескатарите, която е почти 100% растителна и базирана на риба. Също така, не забравяйте да ограничите консумацията на риба до две или три порции всяка седмица, като избягвате риба с по-високо съдържание на живак, като риба тон, риба меч, скумрия и камбала. Ако сте над 65 и започнете да усещате намалена мускулна маса, сила и мазнини, добавете повече риба в диетата си заедно с други храни на животинска основа, включително яйца и специфични сирена, като фета или пекорино, и кисело мляко от козе мляко.
Не яжте твърде много протеин
Според плана за диета за дълголетие, ние трябва да ядем 0.31 на 0.36 грама протеин на килограм телесни мазнини всеки ден. Ако претеглите 130lbs, трябва да ядете около 40 на 47 грама протеин на ден, или еквивалент на 1.5 филе от сьомга, чаша 1 от нахут или чаши 2 1 / 2 от леща, от които 30 грама трябва да се консумират в едно хранене. Ако претеглите 200 до 220lbs, трябва да ядете около 60 на 70 грама протеин на ден, или еквивалент на две филета от сьомга, чашки 3 1 / 2 от леща или чаши 1 1 / 2 от нахут. Консумацията на протеини трябва да се увеличи след 65. За по-голямата част от нас, 10 до 20 процент увеличение или 5 на 10 грама повече всеки ден, е достатъчно. И накрая, диетата за дълголетие е свободна от животински протеини като червено месо, бяло месо и птици, с изключение на животински протеини в рибата. Тази уникална диетична програма е сравнително висока в растителните протеини като бобови и ядки за оптимизиране на здравето и здравето.
Увеличете добрите мазнини и сложните въглехидрати
Като част от плана за хранене на дълголетието, трябва да се консумират по-големи количества полиненаситени мазнини, като тези, които се намират в сьомга, бадеми, орехи и зехтин, докато трябва да се ядат по-малки количества наситени, хидрогенирани и транс-мазнини. По същия начин, като част от плана за диета за дълголетие, вие също трябва да ядете сложни въглехидрати, като тези, които се намират в пълнозърнестия хляб, бобовите растения и зеленчуците. Уверете се, че сте ограничили консумацията на макаронени изделия, ориз, хляб, плодове и плодови сокове, които могат да бъдат превърнати в захари, докато стигнат до червата.
Вземете хранителни добавки
Човешкото тяло се нуждае от протеини, есенциални мастни киселини като омега-3 и омега-6, витамини, минерали и дори захари, за да функционира правилно. Когато приемът на определени хранителни вещества стане твърде нисък, методите за възстановяване, замяна и защита на човешкото тяло могат да забавят или да спрат, като позволят на гъби, бактерии и вируси да причинят щети, които могат да доведат до различни здравни проблеми. Приемайте витаминни и минерални хранителни добавки, особено за омега-3, както Ви е препоръчал Вашият медицински специалист.
Яжте различни храни от вашия Аncestry
За да вземете всички необходими хранителни вещества, които трябва, трябва да ядете голямо разнообразие от храни, но най-добре е да избирате храни, които са често срещани на масата на вашите родители, баби и дядовци и прабаби. Например, в много северно европейски страни, където обикновено се консумира мляко, непоносимостта към лактоза е сравнително рядка, докато лактозната непоносимост е доста често срещана в южните европейски и азиатски страни, където млякото не е било част от конвенционалната диета за възрастни. Ако човек с японски произход, живеещ в Съединените щати, внезапно реши да започне да пие мляко, което вероятно рядко се сервира на масата за хранене на баба и дядо, вероятно ще започнат да се чувстват зле. Най-често срещаните проблеми в тези случаи са непоносимост или автоимунитети, като реакцията на храни, богати на глутен, като хляб и тестени изделия, наблюдавани при хора с цьолиакия. Въпреки че са необходими допълнителни доказателства, е възможно непоносимостта към храни да е свързана с много автоимунни заболявания, включително диабет, колит и болест на Крон.
Яжте две ястия на ден и лека закуска
Според плана за диета за дълголетие е идеално да се яде закуска и едно основно хранене плюс подхранваща нискокалорична закуска с ниско съдържание на захар всеки ден. Докато за някои хора може да се препоръча да ядат три пъти и лека закуска всеки ден. Много хранителни насоки препоръчват да ядем пет до шест хранения всеки ден. Когато хората се съветват да ядат често, често може да им е трудно да регулират приема на калории. През последните двадесет години приблизително 70 процента от населението в САЩ се счита за наднормено тегло или затлъстяване. Много по-трудно е да се преяжда по плана за дълголетие, ако ядете само две и половина хранения всеки ден. Биха били необходими масивни порции бобови растения, зеленчуци и риба, за да се достигне количеството, което би довело до наддаване на тегло. Високото хранене на храната, плюс количеството на храната, изпраща сигнал до стомаха и мозъка ви, че сте имали достатъчно храна. Тази основна система за хранене може понякога да се наложи да бъде разделена на две хранения, за да се избегнат проблеми с храносмилането. Възрастните и възрастните хора, склонни към отслабване, трябва да се хранят три пъти на ден. За хората, които се опитват да отслабнат, както и за хората с наднормено тегло или затлъстяване, най-добрият хранителен съвет би бил да закусват всеки ден; вечеряйте или обядвайте, но не и двете, и заместете пропуснатата храна с една закуска, съдържаща по-малко от 100 калории и не повече от 3 до 5 g захар. Кое хранене пропускате зависи от начина ви на живот, но не се препоръчва да пропуснете закуската поради неблагоприятните здравословни проблеми. Ползата от пропускането на обяда е повече свободно време и енергия. Но има недостатък за ядене на голяма вечеря, особено за хора, които страдат от киселинен рефлукс или проблеми със съня. Недостатъкът на пропускането на вечеря обаче е, че това може да елиминира социалното хранене за деня им.
Яжте в рамките на 12-часов прозорец всеки ден
Друг обичаен хранителен навик, възприет от много столетници, е ограничено във времето за хранене или ограничаване на всички ястия и закуски в рамките на 12-часов прозорец всеки ден. Ефективността на този метод е демонстрирана както в проучвания при хора, така и при животни. Като цяло, вие ще ядете закуска в 8 am и след това вечеряйте с 8 pm. По-кратък прозорец за хранене от десет часа или по-малко може да бъде дори по-добър за загуба на тегло, но е значително по-труден за поддържане и може да увеличи риска от развитие на странични ефекти, като камъни в жлъчката и дори потенциално увеличаване на вероятността от развитие на сърдечно-съдови заболявания. Не трябва да ядете три до четири часа преди сън.
Следвайте диетата за имитация на гладно ProLon
Здравите хора на възраст под 65 трябва да следват Пролонна диета, имитираща гладно, 5-дневна програма за хранене поне два пъти годишно. Шапата е един от ключовите принципи, насърчавани от плана за хранене за дълголетие. Диета, която имитира гладно, предлага същите ползи за здравето от гладно, без действително гладно. Като ядете 800 до 1,100 калории в точни количества и комбинации от храни, които са индивидуално опаковани и етикетирани за всеки ден, можете да „подвеждате” човешкото тяло в състояние на гладно. Чрез различни проучвания, д-р Валтер Лонго открива, че като лишава тялото от храна по този начин, нашите клетки започват да разрушават и регенерират вътрешните ни тъкани, чрез процес, известен като аутофагия, убивайки и замествайки или регенериращи, увредени клетки. Освен това, гладуването може да обърне различни здравни проблеми, да унищожи раковите клетки и значително да намали възможността за развитие на болестта на Алцхаймер.
С плана за диета за дълголетие, представен в книгата на д-р Валтер Лонго, вие ще се храните по-добре, ще се чувствате по-добре и макар да не е създаден като план за отслабване, може дори да свалите няколко килограма. Няма да ви се налага да обмисляте сложни хранителни правила и да правите трудни избори с тази уникална диетична програма. След като разберете тези промени в начина на живот, ще можете да подобрите цялостното си здраве и уелнес вашият дълголетие. - Обхватът на нашата информация е ограничен до хиропрактика, проблеми със здравето на гръбначния стълб и теми за функционална медицина. За по-нататъшно обсъждане на темата, моля не се колебайте да попитате д-р Алекс Хименес или се свържете с нас на адрес 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителна дискусия по темата: Остра болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за увреждане и пропуснати дни по време на работа по целия свят. Болката в гърба се свързва с втората най-често срещана причина за посещения на лекарски кабинет, превъзхождана само от горните дихателни инфекции. Приблизително 80 процента от населението ще изпита болки в гърба поне веднъж през целия си живот. Вашият гръбначен стълб е сложна структура, съставена от кости, стави, сухожилия и мускули сред другите меки тъкани. Наранявания и / или утежнени състояния, като например херния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
КСИМОГЕНИ Ексклузивни професионални формули са достъпни чрез избрани лицензирани професионалисти в областта на здравеопазването. Продажбата по интернет и дисконтирането на формулите XYMOGEN са строго забранени.
Гордост, Д-р Александър Хименес прави формулите XYMOGEN достъпни само за пациенти под нашата грижа.
Моля, обадете се в нашия офис, за да можем да назначим консултация с лекар за незабавен достъп.
Ако сте пациент Медицинска и хиропрактична клиника за нараняване, можете да попитате за XYMOGEN, като се обадите 915-850-0900.
За ваше удобство и преглед на XYMOGEN продукти, моля прегледайте следната връзка. *XYMOGEN-подреждане иИзтегли
* Всички горепосочени политики на XYMOGEN остават в сила.
Инструментът Find A Practitioner на IFM е най-голямата мрежа за препращане във функционалната медицина, създадена, за да помогне на пациентите да намерят практикуващи функционална медицина навсякъде по света. Сертифицираните практикуващи от IFM са изброени на първо място в резултатите от търсенето, като се има предвид тяхното обширно образование по функционална медицина