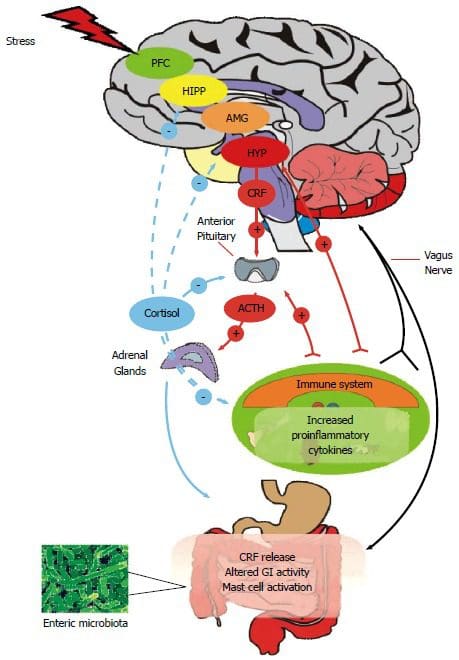
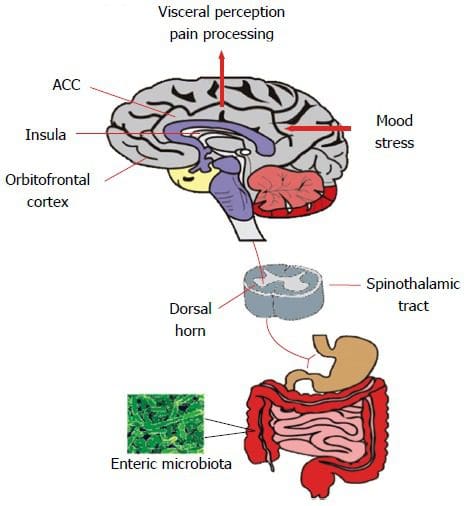
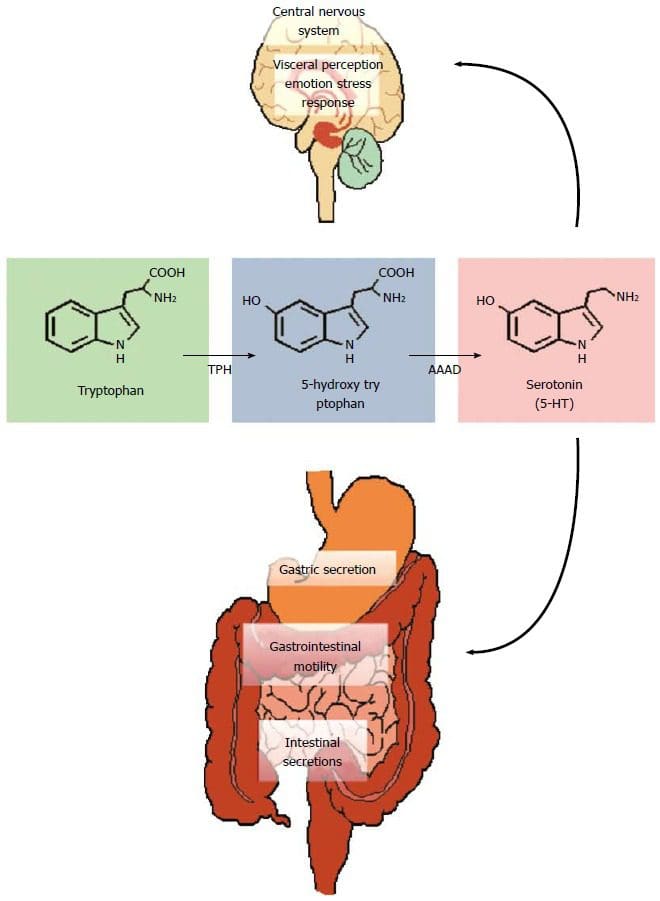
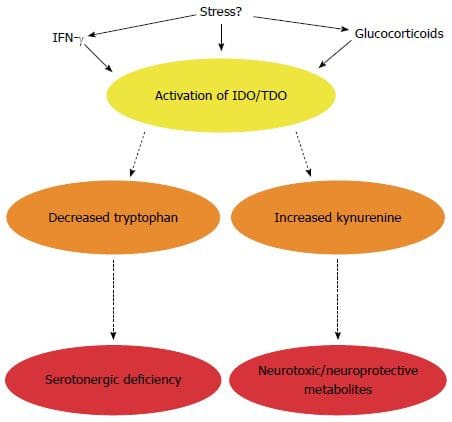
Екип по хиропрактика и функционална медицина на клиниката за оксидативен стрес. Оксидативният стрес се определя като нарушение в баланса между производството на реактивен кислород (свободни радикали) и антиоксидантната защита. С други думи, това е дисбаланс между производството на свободни радикали и способността на организма да противодейства или детоксикира вредните ефекти чрез неутрализиране от антиоксиданти. Оксидативният стрес води до много патофизиологични състояния в организма. Те включват невродегенеративни заболявания, т.е. болестта на Паркинсон, болестта на Алцхаймер, генни мутации, ракови заболявания, синдром на хроничната умора, синдром на крехката Х, заболявания на сърцето и кръвоносните съдове, атеросклероза, сърдечна недостатъчност, инфаркт и възпалителни заболявания. Окислението се случва при редица обстоятелства:
клетките използват глюкоза, за да направят енергия
имунната система се бори с бактериите и създава възпаление
телата детоксикират замърсителите, пестицидите и цигарения дим
Има милиони процеси, които се извършват в тялото ни във всеки един момент, което може да доведе до окисляване. Ето няколко симптома:
Умора
Загуба на памет и мозъчна мъгла
Болка в мускулите и / или ставите
Бръчки заедно със сива коса
Намалено зрение
Главоболие и чувствителност към шума
Податливост към инфекции
Изборът на органични храни и избягването на токсините във вашата среда прави голяма разлика. Това, заедно с намаляването на стреса, може да бъде от полза за намаляване на окисляването.
Да се събудиш уморен дори след получаване на шест или повече часа сън?
Под голямо количество стрес?
Ако изпитвате някоя от тези ситуации, тогава това може да се дължи на нивата на мелатонин и кортизол, засягащи тялото и циркадния ритъм.
В цял свят милиони хора имат проблеми със съня. В Съединените щати има приблизително около 50-70 милиона души които имат лошо качество на съня. Когато човек е спал по-малко от осем часа, той се уморява и много проблеми могат да им дойдат, особено ако животът им е забързан. При забързания начин на живот и лошия сън, това може да накара тялото да има ниска енергия, за да изпълни каквато и да е задача, хормонът на стреса кортизол ще се повиши, а заболявания като високо кръвно налягане и диабет могат да причинят проблеми, които могат да бъдат хронични, ако не са лекувани.
Във функционалната ендокринология мелатонинът и кортизолът са хормони, които тялото произвежда естествено. Кортизолният хормон или стресовият хормон помага на тялото да бъде в състояние на „борба или полет“, което може да бъде добро за всеки, който се занимава с проект или отива на интервю за работа. Въпреки че, когато нивата на кортизоловия хормон са високи, това може да доведе до усложнения като възпаление, хроничен оксидативен стрес и високо кръвно налягане.
Циркадният ритъм на мелатонина
С хормона мелатонин този хормон казва на тялото кога е време да спи. Понякога обаче хората трудно спят и приемането на добавки с мелатонин всъщност може да отпусне тялото и по този начин да накара човек да заспи. Тъй като епифизната жлеза произвежда мелатонин от мозъка, тя може да се намери и в очите, костния мозък и червата, за да се отпусне тялото и да накара човек да заспи по естествен начин. някои проучванията показват че циркадният ритъм на епифизата, която произвежда мелатонин. Правейки това, изследването показва, че прилагането на мелатонин може:
Един: предизвикват съня на хора, които имат проблеми със заспиването.
две: инхибира тялото да се събуди естествено от циркадния пейсмейкър.
Три: преместете циркадните биологични часовници, за да увеличите приема на сън, когато човек се опитва да спи по-рано, за да получи пълните осемчасови ползи от съня.
Когато човек работи на работа от 9 до 5, той се издига с телата си и отпуска телата си след тежък работен ден. Проучванията откриха че хормоните мелатонин и кортизол помагат да се регулира 24-часовият модел на функцията на организма и отговорите му изключително. С цикъла на производство на хормони в организма може да се наруши, ако човекът е буден късно вечер или спи през деня. Когато това се случи, човекът може да получи разрушителни разстройства като промени в настроението, замаяност, да бъде раздразнителен и депресиран и да има метаболитни нарушения. И не само това, но имунната система на организма и неговата ендокринна система също могат да бъдат повредени, което води до приемник на инфекции и болести.
Има повече изследвания за циркадните ритми в тялото, както проучванията показват как хората, които работят в нощната смяна, са били свързани с огромен брой неблагоприятни здравословни проблеми, които атакуват сърдечно-съдовата и стомашно-чревната система, както и нарушават метаболитната система. Всеки, който е работил през нощната смяна, трябва да промени графика си на сън и да се адаптира към бързата преориентация в графика си за сън / събуждане, за да отиде на работа и да си върши работата. Тъй като всички работят по график на смяна, това може да бъде стресиращо и да повлияе на телесната работа на работника, както и да повлияе на секрецията на мелатонин и кортизол.
Начини за подпомагане на кортизола и мелатонина
Изненадващо обаче, има начини за понижаване нивата на кортизола и се уверете, че нивата на мелатонин работят правилно, за да функционира тялото. За да бъдат понижени нивата на кортизола, човек трябва да прави медитативни практики, да открива приятно хоби и най-важното - да опита упражнения за дълбоко дишане, за да отпусне тялото от нежелан стрес. С упражненията за дълбоко дишане може да помогне на тялото да освободи всяко напрежение, което човек държи, а мускулите в тялото започнаха да се отпускат и кръвта започва да тече. С нивата на мелатонин те работят заедно с циркадния ритъм на тялото и се уверяват, че тялото знае кога е време да се събуди, да заспи и да се храни. Хормонът мелатонин също може да помогне за регулиране на телесната температура, т.е. кръвно наляганеи нивата на хормоните, за да се уверите, че функционира правилно. Когато има високи нива на тези системи, това може да накара организма да развие хронични заболявания и да навреди на тялото в процеса.
Проучване показва че мелатониновите хормони могат да се свързват с неврологичните рецептори в тялото, като по този начин насърчават релаксацията. Тъй като мелатонинът се свързва с неврологичните рецептори, той също може да намали нервната активност и нивата на допамин, за да направи очите тежки, като по този начин човекът заспива.
Заключение
Тъй като тялото може да произвежда естествено нивата на мелатонин и кортизол, за да се увери, че тялото не се прекалява със стреса през целия ден. Тъй като мелатонинът е свързан с циркадния ритъм на тялото, тялото знае кога да стои и да заспи. Тъй като всички имат забързан график, е от съществено значение да отделите време и да се отпуснете и да влезете в здравословен график за сън, за да може тялото да бъде здраво и функциониращо. някои продукти са тук, за да се уверим, че ендокринната система функционира правилно и поддържа метаболизма на надбъбречните жлези и захарта.
Обхватът на нашата информация е ограничен до хиропрактика, проблеми с мускулно-скелетното и нервно здраве или статии, теми и дискусии за функционалната медицина. Използваме функционални здравни протоколи за лечение на наранявания или нарушения на опорно-двигателния апарат. Нашият офис направи разумен опит да предостави подкрепящи цитати и определи съответното изследователско проучване или проучвания в подкрепа на нашите публикации. Ние също така правим копия на подкрепящи изследователски проучвания на разположение на съвета и на обществеността при поискване. За да обсъдим допълнително темата по-горе, моля, попитайте д-р Алекс Хименес или се свържете с нас на 915-850-0900.
Литература:
Cajochen, C, et al. „Ролята на мелатонина в регулирането на човешките циркадни ритми и съня“ Вестник по невроендокринология, Национална медицинска библиотека на САЩ, апр. 2003, www.ncbi.nlm.nih.gov/pubmed/12622846.
James, Francine O, et al. „Циркадни ритми на мелатонин, кортизол и експресия на гена на часовника по време на симулирана работа на нощна смяна“. Спя, Associated Professional Sleep Societies, LLC, ноември 2007 г., www.ncbi.nlm.nih.gov/pmc/articles/PMC2082093/.
Monteleone, P, et al. „Временна връзка между мелатонина и кортизола в отговор на физическия стрес през нощта при хората“. Psychoneuroendocrinology, Национална медицинска библиотека на САЩ, 1992, www.ncbi.nlm.nih.gov/pubmed/1609019.
Раман, Райън. „Как мелатонинът може да ви помогне да спите и да се чувствате по-добре“ Healthline, Healthline Media, 3 септември 2017 г., www.healthline.com/nutrition/melatonin-and-sleep.
Zamanian, Zahra, et al. „Схема на промените в циркадните ритми на кортизол и мелатонин в охраната на Университета по медицински науки в Шираз“. Международен вестник за превантивна медицина, Medknow Publications & Media Pvt Ltd, юли 2013 г., www.ncbi.nlm.nih.gov/pmc/articles/PMC3775223/.
Съвременна интегративна и функционална медицина- Есе Куам Видери
Чрез информиране на хората за това как Националният университет по здравни науки предоставя знания за бъдещите поколения, които искат да направят промяна в света. Университетът предлага голямо разнообразие от медицински професии за функционална и интегративна медицина.
Чудили ли сте се защо се чувствате мудни от дълъг ден? Или ви прилошава стомаха, когато сте изяли нещо лошо или сте прекалили с любимата си храна? Възможно ли е червата ви да показват признаци на стрес и дискомфорт поради определени навици, с които може да се сблъскате и дори не сте знаели за това?
В предишната ни статия говорихме за шестте вида храна че червата ни трябва да са здрави. От нашите черва съдържа трилиони микробиоми, както добри, така и лоши, тези микробиоми играят важна роля за цялостното ни здраве. Здравият микробиом подобрява нашата здравето на червата, здравето на сърцето, здравето на мозъка, контролира теглото ни намлява регулира кръвната ни захар. С добрите бактерии в червата ни, бактериите ни са полезни с добра храносмилателна система и унищожават вредните бактерии. Но някои начини на живот и избор на диета всъщност могат да увеличат лошите бактерии и да намалят добрите бактерии и цялостното здраве.
Ето пет изненадващо начина на живот, които нараняват червата ви:
Не ядете широка гама от храни
Нашите черва играят важна роля за цялостното ни здраве. Когато ядем добри пълноценни храни, червата ни са по-щастливи; имаме повече енергия да изпълним всяка задача, която ни е хвърлена и получаваме хранителни вещества за нашата чревна флора. Въпреки това, през последните няколко десетилетия, ние се насочваме повече към преработените храни поради икономическия натиск от увеличеното производство на храни. FOA заяви, че „75 процента от храната в света се произвежда само от 12 растения и пет животински вида“ и това е много лошо за нашата чревна флора.
Тук, в клиника за медицинска и хиропрактика на травми, ние информираме нашите пациенти за важността на храненето на питателни, пълноценни храни за насърчаване не само на здрави черва, но и на здрав ум. Когато тялото се запознае с a голямо разнообразие от пълноценни храни (с високо съдържание на фибри), червата ни започват да поправят щетите от преработената храна, която може да сме консумирали вътрешно.
Въпреки това, когато пренебрегнете пребиотиците към вашата диета, вие сте вреди на храносмилателното ви здраве. Без пребиотици нашата храносмилателна система забавя развитието и разнообразието на нашата чревна флора. Така че, за да имате a здрав микробиом развитие, трябва да включите в диетата си храни, пълни както с смилаеми, така и с несмилаеми фибри. Някои храни, включени в тази категория, са овес, ядки, лук, чесън, праз, аспержи, банани, круши, нахут и боб.
Придържането към диета с високо съдържание на фибри може би е предизвикателство, но има възможност за приемане на пребиотични добавки. Ако имате хранителен алерген или хранителна чувствителност към храни с високо съдържание на фибри, приемайте пребиотични добавки всъщност може да помогне за растежа на Bifidobacterium и Faecalibacterium в червата ви и да бъде от полза за вашето здраве без дискомфорт.
Прекомерна консумация на алкохол
Всеки възрастен се наслаждава на алкохола от време на време. Да, това е една от онези напитки, които ви помагат да се отпуснете малко след дълъг ден, но твърде много от тях може да доведе до злоупотреба с алкохол и пристрастяване. И така, знаете ли, че консумацията на толкова много алкохол е вредна вашето сърце, черен дроб и мозък; по този начин наранява здравето на червата и ви причинява дисбиоза?
Едно проучване заяви, че алкохолиците с дисбиоза имат по-ниско средно изобилие от Bacteroidetes и голямо изобилие от Proteobacteria. Тези, които не са били алкохолици, не са били засегнати от проучването.
Въпреки това; има някои добри новини за ограничаването на алкохолизма и че това може да бъде полезно за чревните бактерии. Ако сте консумирали умерено отговорно червено вино, полифеноли във виното може да помогне на вашата чревна флора. Така че, насладете се на чаша вино от време на време като малко лакомство, което не трябва да се приема за даденост.
Неадекватен сън
В една от предишните статии говорихме как да постигнем a добър нощен сън чрез билки. Когато почти не спим през нашия забързан живот, това ни засяга чрез различни здравословни проблеми, включително сърдечно заболяване намлява затлъстяване, В 2016 проучване, изследователите откриха ефекта от краткосрочното лишаване от сън върху чревната микробиота след два дни.
Когато тялото ни не получава препоръчителните 8 часа сън, червата ни поемат огромно влияние, тъй като се чувстваме бавни и изтощени. Така че, за да сме сигурни, че нашите микробиома на червата ще бъдат погрижени, препоръчваме ви да изключите електронните си устройства поне 30 минути, преди да се приготвите да се настаните за през нощта. Изключете всички светлини и не пийте никакви течности поне два часа преди лягане, затворете очи и поемете дълбоко въздух в медитативно състояние и се отпуснете, докато се унасяте в спящия град.
Неадекватни упражнения
Поради нашия забързан начин на живот и стресираща работа е трудно да намерим време за упражнения. Но когато наистина намираме време за упражнения, умът ни не само се чувства добре; но тялото и червата ни също се чувстват добре. Въпреки това, нещата винаги изникват, когато сме в рутинна тренировка и трябва да пропуснем упражненията напълно. Случва се на всички ни и е трудно да продължим откъдето сме спрели, когато се опитахме да тренираме.
Когато не тренираме поне няколко пъти в седмицата, телата ни оказват огромно влияние върху нас, докато напълняваме, нашите стресът е твърде висок, и имаме a по-голям шанс за получаване на хронично заболяване. Когато това се случи, нашата чревна флора е огромен недостатък. Тук в клиниката се стремим да информираме нашите пациенти за важността на упражненията и че те не само променят живота им, но и променят изцяло настроението им.
Въпреки това, не влизайте само в тежки рутинни упражнения, при които ще се нараните. Започнете с тренировка с ниска интензивност, след което я надградете, докато вървите, защото чревната ви флора ще ви благодари за това.
Като последна дума, ние тук от Injury Medical искаме да ви информираме за храненето и начините да ви помогнем да подобрите заболяванията си с тези 5 изненади. Но също така да ви образова какво може да нарани червата ви. С тези изненади и леки промени в ежедневието ви, червата ви ще ви благодарят за дългия път.
Ресурси на НЦБИ
Според данни от изследване на 2016, имунната система на червата е от основно значение за предотвратяване на различни заболявания и често може да допринесе за метаболитни нарушения. Въпреки това, това може също да помогне за постигане на цел на лечението, когато се наблюдава системно възпаление при инсулинова резистентност. Освен това, модифицираният чревен имунитет е свързан с промени в чревната микробиота, функцията на чревната бариера, имунните клетки в червата и резистентността към антигени, които влизат в стомашно-чревната или стомашно-чревната система. Въпреки че преди се е смятало, че това повишава опасността от заболявания на хранопровода, включително патогенни инфекции и хронично възпаление, което в крайна сметка може да доведе до хронични здравословни проблеми.
Кетонните тела се създават от черния дроб и се използват като източник на енергия, когато глюкозата не е лесно достъпна в човешкото тяло. Двете основни кетонни тела са ацетоацетат (AcAc) и 3-бета-хидроксибутират (3HB), докато ацетонът е третото и най-малко разпространено кетонно тяло. Кетоните винаги присъстват в кръвта и нивата им се повишават по време на гладуване и продължителни упражнения. Кетогенеза е биохимичният процес, чрез който организмите произвеждат кетонни тела чрез разграждането на мастни киселини и кетогенни аминокиселини.
Кетонните тела се генерират главно в митохондриите на чернодробните клетки. Кетогенезата възниква, когато има ниски нива на глюкоза в кръвта, особено след изчерпване на други клетъчни запаси от въглехидрати, като гликоген. Този механизъм може да възникне и при недостатъчни количества инсулин. Производството на кетонни тела в крайна сметка започва, за да направи достъпна енергия, която се съхранява в човешкото тяло като мастни киселини. Кетогенезата се случва в митохондриите, където се регулира независимо.
абстрактен
Метаболизмът на кетонните тела е централен възел във физиологичната хомеостаза. В този преглед ние обсъждаме как кетоните служат на отделни метаболитни роли за фина настройка, които оптимизират работата на органите и организма в различни остатъци от хранителни вещества и предпазват от възпаление и нараняване в множество органни системи. Традиционно разглеждани като метаболитни субстрати, включени само при ограничаване на въглехидратите, последните наблюдения подчертават важността на кетонните тела като жизненоважни метаболитни и сигнални медиатори, когато въглехидратите са в изобилие. Допълвайки репертоара от известни терапевтични възможности за заболявания на нервната система, се появиха перспективни роли на кетонните тела при рак, както и интригуващи защитни роли в сърцето и черния дроб, отваряйки терапевтични възможности при свързани със затлъстяването и сърдечно-съдови заболявания. Обсъждат се противоречия в кетонния метаболизъм и сигнализирането, за да се съгласуват класическата догма със съвременните наблюдения.
Въведение
Кетонните тела са жизненоважен алтернативен метаболитен източник на гориво за всички области на живота, еукария, бактерии и археи (Aneja et al., 2002; Cahill GF Jr, 2006; Krishnakumar et al., 2008). Метаболизмът на кетонните тела при хората е използван за подхранване на мозъка по време на епизодични периоди на недостиг на хранителни вещества. Кетонните тела са преплетени с важни метаболитни пътища при бозайниците като ?-окисление (FAO), цикъл на трикарбоксилната киселина (TCA), глюконеогенеза, de novo липогенеза (DNL) и биосинтеза на стероли. При бозайниците кетонните тела се произвеждат предимно в черния дроб от ацетил-КоА, извлечен от FAO, и се транспортират до екстрахепаталните тъкани за терминално окисление. Тази физиология осигурява алтернативно гориво, което се допълва от сравнително кратки периоди на гладуване, което увеличава наличността на мастни киселини и намалява наличността на въглехидрати (Cahill GF Jr, 2006; McGarry and Foster, 1980; Robinson and Williamson, 1980). Окислението на кетонните тела става значителен принос за цялостния енергиен метаболизъм на бозайниците в извънчернодробните тъкани в безброй физиологични състояния, включително гладуване, гладуване, неонаталния период, след тренировка, бременност и придържане към диети с ниско съдържание на въглехидрати. Концентрациите на циркулиращите общи кетонни тела при здрави възрастни хора обикновено показват циркадианни колебания между приблизително 100-250 µM, повишават се до ~1 mM след продължително упражнение или 24 часа гладуване и могат да се натрупват до 20 mM при патологични състояния като диабетна кетоацидоза ( Cahill GF Jr, 2006; Johnson et al., 1969b; Koeslag et al., 1980; Robinson and Williamson, 1980; Wildenhoff et al., 1974). Човешкият черен дроб произвежда до 300 g кетонни тела на ден (Balasse and Fery, 1989), които допринасят между 5% от общия разход на енергия в нахранени, гладни и гладни състояния (Balasse et al., 20; Cox et al. др., 1978).
Последните проучвания подчертават императивната роля на кетонните тела в метаболизма на клетките на бозайниците, хомеостазата и сигнализирането при голямо разнообразие от физиологични и патологични състояния. Освен че служат като енергийни горива за извънчернодробни тъкани като мозък, сърце или скелетни мускули, кетонните тела играят основна роля като сигнални медиатори, двигатели на протеинова пост-транслационна модификация (PTM) и модулатори на възпаление и оксидативен стрес. В този преглед ние предоставяме както класически, така и съвременни възгледи за плейотропните роли на кетонните тела и техния метаболизъм.
Преглед на метаболизма на кетонното тяло
Скоростта на чернодробната кетогенеза се управлява от организирана серия от физиологични и биохимични трансформации на мазнините. Първичните регулатори включват липолиза на мастни киселини от триацилглицероли, транспорт до и през плазмената мембрана на хепатоцитите, транспорт в митохондриите чрез карнитин палмитоилтрансфераза 1 (CPT1), β-окислителна спирала, активност на TCA цикъла и междинни концентрации, потенциал на редокс регулатор и хормонален регулаторен потенциал и хормонален регулаторен ефект. от тези процеси, предимно глюкагон и инсулин [прегледани в (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al., 1983; Kahn et al., 2005; McGarry and Foster , 1980; Williamson et al., 1969)]. Класически кетогенезата се разглежда като преливащ път, при който ацетил-КоА, получен от а-окисление, превишава активността на цитрат синтазата и/или наличността на оксалоацетат за кондензация за образуване на цитрат. Междинните съединения с три въглерода проявяват антикетогенна активност, вероятно поради способността им да разширяват оксалоацетатния пул за консумация на ацетил-CoA, но концентрацията на ацетил-CoA в черния дроб сама по себе си не определя кетогенната скорост (Foster, 1967; Rawat and Menahan, 1975; Williamson et al., 1969). Регулирането на кетогенезата чрез хормонални, транскрипционни и пост-транслационни събития заедно подкрепят идеята, че молекулярните механизми, които прецизират кетогенната скорост, остават ненапълно разбрани (вижте Регулация на HMGCS2 и SCOT/OXCT1).
Кетогенезата се извършва предимно в чернодробния митохондриален матрикс със скорост, пропорционална на общото окисление на мазнините. След транспортиране на ацилови вериги през митохондриалните мембрани и ?-окисление, митохондриалната изоформа на 3-хидроксиметилглутарил-CoA синтаза (HMGCS2) катализира съдбата, извършвайки кондензация на ацетоацетил-CoA (AcAc-CoA) и генериране на HMG-Ac-CoA (фиг. 1А). HMG-CoA лиаза (HMGCL) разцепва HMG-CoA, за да освободи ацетил-CoA и ацетоацетат (AcAc), а последният се редуцира до d-p-хидроксибутират (d-pOHB) от фосфатидилхолин-зависима митохондриална d-pOHB дехидрогеназа (D-pOHB дехидрогеназа). BDH1) в NAD+/NADH-свързана почти равновесна реакция (Bock and Fleischer, 1975; LEHNINGER et al., 1960). Равновесната константа на BDH1 благоприятства производството на d-?OHB, но съотношението на AcAc/d-?OHB кетонни тела е право пропорционално на съотношението NAD+/NADH на митохондриите и по този начин активността на BDH1 оксидоредуктазата модулира митохондриалния редокс потенциал (Krebs et al., 1969; Williamson et al., 1967). AcAc може също така спонтанно да се декарбоксилира до ацетон (Pedersen, 1929), източник на сладка миризма при хора, страдащи от кетоацидоза (т.е. общи серумни кетонни тела > ~7 mM; AcAc pKa 3.6, ?OHB pKa 4.7). Механизмите, чрез които кетонните тела се транспортират през вътрешната мембрана на митохондриите, не са известни, но AcAc/d-?OHB се освобождават от клетките чрез монокарбоксилатни транспортери (при бозайници, MCT 1 и 2, известни също като носители на разтворено вещество 16A членове 1 и 7) и транспортирани в циркулацията до екстрахепаталните тъкани за терминално окисление (Cotter et al., 2011; Halestrap and Wilson, 2012; Halestrap, 2012; Hugo et al., 2012). Концентрациите на циркулиращи кетонни тела са по-високи от тези в екстрахепаталните тъкани (Harrison and Long, 1940), което показва, че кетонните тела се транспортират надолу по градиент на концентрацията. Мутациите със загуба на функция в MCT1 са свързани със спонтанни пристъпи на кетоацидоза, което предполага критична роля във вноса на кетонни тела.
С изключение на потенциалното отклоняване на кетонните тела в неокислителни съдби (вижте Неокислителни метаболитни съдби на кетонните тела), хепатоцитите нямат способността да метаболизират кетонните тела, които произвеждат. Кетонните тела, синтезирани de novo от черния дроб, (i) се катаболизират в митохондриите на екстрахепаталните тъкани до ацетил-CoA, който е наличен в TCA цикъла за терминално окисление (фиг. 1A), (ii) се отклонява към пътищата на липогенезата или синтеза на стерол ( Фиг. 1В), или (iii) се екскретира с урината. Като алтернативно енергийно гориво, кетонните тела се окисляват жадно в сърцето, скелетните мускули и мозъка (Balasse and Fery, 1989; Bentourkia et al., 2009; Owen et al., 1967; Reichard et al., 1974; Sultan, 1988 ). Екстрахепаталната митохондриална BDH1 катализира първата реакция на окисление на ?OHB, превръщайки я в обратно AcAc (LEHNINGER et al., 1960; Sandermann et al., 1986). Цитоплазмената d-?OHB-дехидрогеназа (BDH2) само с 20% идентичност на последователността с BDH1 има висок Km за кетонни тела и също така играе роля в хомеостазата на желязо (Davuluri et al., 2016; Guo et al., 2006) . В екстрахепаталния митохондриален матрикс, AcAc се активира до AcAc-CoA чрез обмен на CoA-част от сукцинил-CoA в реакция, катализирана от уникална CoA трансфераза на бозайници, сукцинил-CoA:3-оксокиселина-CoA трансфераза (SCOT, CoA трансфераза; кодиран от OXCT1), чрез реакция, близка до равновесие. Свободната енергия, освободена от хидролизата на AcAc-CoA, е по-голяма от тази на сукцинил-CoA, което благоприятства образуването на AcAc. По този начин окислителният поток на кетонните тела възниква поради масово действие: изобилното снабдяване с AcAc и бързата консумация на ацетил-CoA чрез цитрат синтаза благоприятства образуването на AcAc-CoA (+ сукцинат) от SCOT. По-специално, за разлика от глюкозата (хексокиназа) и мастните киселини (ацил-КоА синтетази), активирането на кетонните тела (SCOT) в окисляема форма не изисква инвестиция на АТФ. Обратима AcAc-CoA тиолазна реакция [катализирана от всяка от четирите митохондриални тиолази, кодирани от ACAA2 (кодиращ ензим, известен като T1 или CT), ACAT1 (кодиращ T2), HADHA или HADHB] дава две молекули ацетил-CoA които влизат в TCA цикъла (Hersh and Jencks, 1967; Stern et al., 1956; Williamson et al., 1971). По време на кетозни състояния (т.е. общи серумни кетони > 500 M), кетонните тела допринасят значително за разхода на енергия – и се използват в тъканите бързо, докато настъпи поглъщане или насищане с окисляване (Balasse et al., 1978; Balasse and Fery, 1989 Edmond et al., 1987). Много малка част от извлечените от черния дроб кетонни тела могат лесно да бъдат измерени в урината, а степента на използване и реабсорбция от бъбреците са пропорционални на циркулиращата концентрация (Goldstein, 1987; Robinson and Williamson, 1980). По време на силно кетотични състояния (> 1 mM в плазмата), кетонурията служи като полуколичествен репортер на кетоза, въпреки че повечето клинични анализи на кетонни тела в урината откриват AcAc, но не и ?OHB (Klocker et al., 2013).
Кетогенни субстрати и тяхното влияние върху метаболизма на хепатоцитите
Кетогенните субстрати включват мастни киселини и аминокиселини (фиг. 1В). Катаболизмът на аминокиселините, особено на левцина, генерира около 4% от кетонните тела в постабсорбционно състояние (Thomas et al., 1982). Така ацетил-CoA субстратният пул за генериране на кетонни тела произлиза главно от мастни киселини, тъй като по време на състояния на намалено снабдяване с въглехидрати, пируватът влиза в чернодробния TCA цикъл предимно чрез анаплероза, т.е. АТФ-зависимо карбоксилиране до оксалоацетат (OAA) или до малат (MAL), а не окислително декарбоксилиране до ацетил-КоА (Jeoung et al., 2012; Magnusson et al., 1991; Merritt et al., 2011). В черния дроб глюкозата и пируватът допринасят незначително за кетогенезата, дори когато декарбоксилирането на пируват до ацетил-КоА е максимално (Jeoung et al., 2012).
Ацетил-КоА включва няколко роли, които са неразделна част от чернодробния междинен метаболизъм извън генерирането на АТФ чрез терминално окисление (вижте също Интегрирането на метаболизма на кетоновите тела, пост-транслационната модификация и клетъчната физиология). Ацетил-КоА алостерично активира (i) пируват карбоксилаза (PC), като по този начин активира механизъм за метаболитен контрол, който увеличава анаплеротично навлизане на метаболити в TCA цикъла (Owen et al., 2002; Scrutton and Utter, 1967) и (ii) дехидрогеназа киназа, която фосфорилира и инхибира пируват дехидрогеназа (PDH) (Cooper et al., 1975), като по този начин допълнително засилва потока на пируват в TCA цикъла чрез анаплероза. Освен това, цитоплазменият ацетил-CoA, чийто пул се увеличава от механизми, които превръщат митохондриалния ацетил-CoA в транспортируеми метаболити, инхибира окисляването на мастни киселини: ацетил-CoA карбоксилазата (ACC) катализира превръщането на ацетил-CoA в малонил-CoA, липогенния субстрат и алостеричен инхибитор на митохондриален CPT1 [прегледан в (Kahn et al., 2005; McGarry and Foster, 1980)]. По този начин, митохондриалният пул ацетил-КоА едновременно регулира и се регулира от преливащия път на кетогенезата, който ръководи ключови аспекти на чернодробния междинен метаболизъм.
Неокислителни метаболитни съдби на кетонните тела
Доминиращата съдба на кетони, получени от черния дроб, е SCOT-зависимо екстрахепатално окисление. Въпреки това, AcAc може да бъде изнесен от митохондриите и използван в анаболни пътища чрез превръщане в AcAc-CoA чрез ATP-зависима реакция, катализирана от цитоплазмената ацетоацетил-CoA синтетаза (AACS, Фиг. 1B). Този път е активен по време на развитието на мозъка и в кърмещата млечна жлеза (Morris, 2005; Robinson and Williamson, 1978; Ohgami et al., 2003). AACS също е силно експресиран в мастната тъкан и активираните остеокласти (Aguilo et al., 2010; Yamasaki et al., 2016). Цитоплазмената AcAc-CoA може да бъде или насочена от цитозолен HMGCS1 към биосинтеза на стерол, или разцепена от една от двете цитоплазмени тиолази до ацетил-CoA (ACAA1 и ACAT2), карбоксилирана до малонил-CoA и да допринесе за синтеза на мастни киселини и др. al., 1984; Edmond, 1974; Endemann et al., 1982; Geelen et al., 1983; Webber и Edmond, 1977).
Докато физиологичното значение все още не е установено, кетоните могат да служат като анаболни субстрати дори в черния дроб. В изкуствен експериментален контекст, AcAc може да допринесе за до половината от новосинтезирания липид и до 75% от новия синтезиран холестерол (Endemann et al., 1982; Geelen et al., 1983; Freed et al., 1988). Тъй като AcAc се получава от непълно окисление на чернодробните мазнини, способността на AcAc да допринесе за липогенезата in vivo би означавало безполезен чернодробен цикъл, при който кетони, получени от мазнини, могат да бъдат използвани за производството на липиди, понятие, чието физиологично значение изисква експериментално валидиране, но може да служи адаптивни или неадаптивни роли (Solinas et al., 2015). AcAc жадно осигурява холестерогенеза, с нисък AACS Km-AcAc (~50 µM), благоприятстващ активирането на AcAc дори в състояние на хранене (Bergstrom et al., 1984). Предполага се, че динамичната роля на метаболизма на цитоплазмения кетони в първичните миши ембрионални неврони и в адипоцитите, получени от 3T3-L1, тъй като нокдаунът на AACS нарушава диференциацията на всеки клетъчен тип (Hasegawa et al., 2012a; Hasegawa et al., ). Нокдаунът на AACS при мишки in vivo намалява серумния холестерол (Hasegawa et al., 2012c). SREBP-2012, главен транскрипционен регулатор на биосинтезата на холестерола и рецептор, активиран от пероксизомен пролифератор (PPAR)-? са AACS транскрипционни активатори и регулират транскрипцията му по време на развитието на неврит и в черния дроб (Aguilo et al., 2; Hasegawa et al., 2010c). Взети заедно, метаболизмът на цитоплазменото кетонно тяло може да бъде важен при избрани състояния или естествена история на заболяването, но е недостатъчен за изхвърляне на кетонни тела, получени от черния дроб, тъй като масивна хиперкетонемия възниква в условията на селективно увреждане на първичната окислителна съдба чрез загуба на функционални мутации до SCOT (Berry et al., 2012; Cotter et al., 2001).
Регулиране на HMGCS2 и SCOT/OXCT1
Отклонението на митохондриално от гена, кодиращ цитозолния HMGCS, се е случило в началото на еволюцията на гръбначните, поради необходимостта от подкрепа на чернодробната кетогенеза при видове с по-високо съотношение на мозъка към телесното тегло (Boukaftane et al., 1994; Cunnane и Crawford, 2003). Естествено срещащите се HMGCS2 мутации със загуба на функция при хора причиняват пристъпи на хипокетотична хипогликемия (Pitt et al., 2015; Thompson et al., 1997). Здравата експресия на HMGCS2 е ограничена до хепатоцитите и епитела на дебелото черво и нейната експресия и ензимната активност се координират чрез различни механизми (Mascaro et al., 1995; McGarry and Foster, 1980; Robinson and Williamson, 1980). Докато пълният обхват на физиологичните състояния, които влияят на HMGCS2, изисква допълнително изясняване, неговата експресия и/или активност се регулират по време на ранния постнатален период, стареене, диабет, гладуване или поглъщане на кетогенна диета (Balasse and Fery, 1989; Cahill GF Jr, 2006 ; Girard et al., 1992; Hegardt, 1999; Satapati et al., 2012; Sengupta et al., 2010). При плода метилирането на 5 фланкираща област на гена Hmgcs2 обратно корелира с неговата транскрипция и е частично обърнато след раждането (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al. ., 1983). По подобен начин, чернодробният Bdh1 проявява модел на експресия на развитие, нарастващ от раждането до отбиването и също се индуцира от кетогенна диета по начин, зависим от фибробластния растежен фактор (FGF)-21 (Badman et al., 2007; Zhang et al., 1989 ). Кетогенезата при бозайници е силно реагираща както на инсулин, така и на глюкагон, като съответно се потиска и стимулира (McGarry and Foster, 1977). Инсулинът потиска липолизата на мастната тъкан, като по този начин лишава кетогенезата от нейния субстрат, докато глюкагонът увеличава кетогенния поток чрез директен ефект върху черния дроб (Hegardt, 1999). Транскрипцията на Hmgcs2 се стимулира от транскрипционния фактор на вилицата FOXA2, който се инхибира чрез инсулин-фосфатидилинозитол-3-киназа/Akt и се индуцира от глюкагон-cAMP-p300 сигнализиране (Arias et al., 1995; Hegardt, 1999; , 1990; Thumelin et al., 1993; von Meyenn et al., 2013; Wolfrum et al., 2004; Wolfrum et al., 2003). PPAR? (Rodriguez et al., 1994) заедно с целта си, FGF21 (Badman et al., 2007) също индуцира транскрипция на Hmgcs2 в черния дроб по време на гладуване или прилагане на кетогенна диета (Badman et al., 2007; Inagaki et al., 2007 ). Индукция на PPAR? може да се случи преди прехода от фетална към неонатална физиология, докато активирането на FGF21 може да бъде благоприятно в ранния неонатален период чрез ?OHB-медиирано инхибиране на хистон деацетилаза (HDAC)-3 (Rando et al., 2016). mTORC1 (мишена на рапамицин комплекс 1 при бозайници) зависимо инхибиране на PPAR? транскрипционната активност също е ключов регулатор на генната експресия на Hmgcs2 (Sengupta et al., 2010), а чернодробният PER2, главен циркаден осцилатор, индиректно регулира експресията на Hmgcs2 (Chavan et al., 2016). Последните наблюдения показват, че екстрахепаталният тумор-индуциран интерлевкин-6 уврежда кетогенезата чрез PPAR? потискане (Flint et al., 2016).
HMGCS2 ензимната активност се регулира чрез множество PTM. Фосфорилирането на серин на HMGCS2 повишава неговата активност in vitro (Grimsrud et al., 2012). Активността на HMGCS2 се инхибира алостерично чрез сукцинилиране на сукцинил-CoA и лизинов остатък (Arias et al., 1995; Hegardt, 1999; Lowe and Tubbs, 1985; Quant et al., 1990; Rardin et al., 2013; 1975; Thumelin et al., 1993). Сукцинилирането на HMGCS2, HMGCL и BDH1 лизинови остатъци в чернодробните митохондрии са мишени на NAD+ зависимата деацилаза сиртуин 5 (SIRT5) (Rardin et al., 2013). Активността на HMGCS2 също се засилва от деацетилиране на SIRT3 лизин и е възможно кръстосаните смущения между ацетилирането и сукцинилирането да регулират активността на HMGCS2 (Rardin et al., 2013; Shimazu et al., 2013). Въпреки способността на тези PTM да регулират HMGCS2 Km и Vmax, флуктуациите на тези PTM все още не са внимателно картографирани и не са потвърдени като механистични двигатели на кетогенезата in vivo.
SCOT се експресира във всички клетки на бозайници, които съдържат митохондрии, с изключение на тези на хепатоцитите. Значението на активността на SCOT и кетолизата е демонстрирано при мишки SCOT-KO, които показват еднаква смъртност поради хиперкетонемична хипогликемия в рамките на 48 часа след раждането (Cotter et al., 2011). Тъканно-специфичната загуба на SCOT в неврони или скелетни миоцити предизвиква метаболитни аномалии по време на гладуване, но не е смъртоносна (Cotter et al., 2013b). При хората дефицитът на SCOT се проявява в началото на живота с тежка кетоацидоза, причиняваща летаргия, повръщане и кома (Berry et al., 2001; Fukao et al., 2000; Kassovska-Bratinova et al., 1996; Niezen-Koning et al. , 1997; Saudubray et al., 1987; Snyderman et al., 1998; Tildon и Cornblath, 1972). Относително малко се знае на клетъчно ниво за SCOT гена и регулаторите на експресия на протеини. Експресията на Oxct1 иРНК и SCOT протеинът и активността са намалени в кетозни състояния, вероятно чрез PPAR-зависими механизми (Fenselau and Wallis, 1974; Fenselau and Wallis, 1976; Grinblat et al., 1986; Okuda et al., 1991; Turko et al. ., 2001; Wentz et al., 2010). При диабетна кетоацидоза несъответствието между чернодробната кетогенеза и екстрахепаталното окисление се влошава от увреждане на активността на SCOT. Свръхекспресията на инсулин-независим глюкозен транспортер (GLUT1/SLC2A1) в кардиомиоцитите също инхибира експресията на Oxct1 ген и регулира надолу терминалното окисляване на кетоните в некетотично състояние (Yan et al., 2009). В черния дроб изобилието на Oxct1 иРНК се потиска от микроРНК-122 и метилиране на хистон H3K27me3, които са очевидни по време на прехода от фетален към неонатален период (Thorrez et al., 2011). Въпреки това, потискането на чернодробната експресия на Oxct1 в постнаталния период се дължи главно на евакуацията на Oxct1-експресиращи хематопоетични прогенитори от черния дроб, а не на загуба на съществуваща преди това експресия на Oxct1 в терминално диференцирани хепатоцити. Всъщност експресията на Oxct1 mRNA и SCOT протеин в диференцирани хепатоцити е изключително ниска (Orii et al., 2008).
SCOT също се регулира от PTM. Ензимът е хипер-ацетилиран в мозъците на SIRT3 KO мишки, които също показват намалено AcAc зависимо производство на ацетил-CoA (Dittenhafer-Reed et al., 2015). Неензимното нитриране на тирозинови остатъци на SCOT също отслабва неговата активност, което е докладвано в сърцата на различни модели на диабетни мишки (Marcondes et al., 2001; Turko et al., 2001; Wang et al., 2010a). За разлика от това, нитрирането на триптофановите остатъци увеличава активността на SCOT (Br g re et al., 2010; Rebrin et al., 2007). Молекулни механизми на специфично за остатъци нитриране или денитриране, предназначени да модулират активността на SCOT, могат да съществуват и изискват изясняване.
Противоречия в екстрахепаталната кетогенеза
При бозайниците основният кетогенен орган е черният дроб и само хепатоцитите и чревните епителни клетки в изобилие експресират митохондриалната изоформа на HMGCS2 (Cotter et al., 2013a; Cotter et al., 2014; McGarry and Foster, 1980; Robinson1980 и Williamson) . Анаеробната бактериална ферментация на сложни полизахариди води до бутират, който се абсорбира от колоноцитите при бозайници за терминално окисление или кетогенеза (Cherbuy et al., 1995), което може да играе роля в диференциацията на колоноцитите (Wang et al., 2016). С изключение на чревните епителни клетки и хепатоцити, HMGCS2 почти липсва в почти всички други клетки на бозайници, но перспективата за екстрахепатална кетогенеза е повишена в туморните клетки, астроцитите на централната нервна система, бъбреците, панкреаса ? клетки, пигментен епител на ретината (RPE) и дори в скелетните мускули (Adijanto et al., 2014; Avogaro et al., 1992; El Azzouny et al., 2016; Grabacka et al., 2016; Kang et al., 2015 ; Le Foll et al., 2014; Nonaka et al., 2016; Takagi et al., 2016a; Thevenet et al., 2016; Zhang et al., 2011). Ектопичен HMGCS2 е наблюдаван в тъкани, които нямат нетен кетогенен капацитет (Cook et al., 2016; Wentz et al., 2010), а HMGCS2 проявява проспективни независими от кетогенезата „моунлайтни“ активности, включително в рамките на клетъчното ядро (Chen et al. , 2016; Костюк и др., 2010; Меертенс и др., 1998).
Всяка екстрахепатална тъкан, която окислява кетонни тела, също има потенциал да натрупва кетонни тела чрез HMGCS2 независими механизми (фиг. 2А). Въпреки това, няма екстрахепатална тъкан, в която концентрацията на кетонни тела в стационарно състояние да надвишава тази в кръвообращението (Cotter et al., 2011; Cotter et al., 2013b; Harrison and Long, 1940), подчертавайки, че кетонните тела се транспортират надолу. концентрационен градиент чрез MCT1/2-зависими механизми. Един механизъм на очевидна екстрахепатална кетогенеза може всъщност да отразява относително увреждане на окислението на кетон. Допълнителни потенциални обяснения попадат в сферата на образуването на кетонни тела. Първо, de novo кетогенезата може да се случи чрез обратима ензимна активност на тиолаза и SCOT (Weidemann and Krebs, 1969). Когато концентрацията на ацетил-КоА е относително висока, реакциите, които обикновено са отговорни за окисляването на AcAc, протичат в обратна посока (GOLDMAN, 1954). Втори механизъм възниква, когато междинните продукти, получени от α-окисление, се натрупват поради затруднено място в цикъла на TCA, AcAc-CoA се превръща в l-?OHB-CoA чрез реакция, катализирана от митохондриална 3-хидроксиацил-CoA дехидрогеназа и допълнително от 3-хидроксибутирил CoA деацилаза до l-?OHB, която е неразличима чрез масспектрометрия или резонансна спектроскопия от физиологичния енантиомер d-?OHB (Reed and Ozand, 1980). l-?OHB може да бъде хроматографски или ензимно разграничен от d-?OHB и присъства в екстрахепаталните тъкани, но не и в черния дроб или кръвта (Hsu et al., 2011). Чернодробната кетогенеза произвежда само d-?OHB, единственият енантиомер, който е BDH субстрат (Ito et al., 1984; Lincoln et al., 1987; Reed and Ozand, 1980; Scofield et al., 1982; Scofield et al., 1982 г.). Трети HMGCS2-независим механизъм генерира d-?OHB чрез катаболизъм на аминокиселини, особено този на левцин и лизин. Четвъртият механизъм е само привиден, защото се дължи на артефакт на етикетиране и по този начин се нарича псевдокетогенеза. Това явление се дължи на обратимостта на реакциите на SCOT и тиолазата и може да причини надценяване на оборота на кетонните тела поради изотопното разреждане на маркера на кетонните тела в екстрахепаталната тъкан (Des Rosiers et al., 1990; Fink et al., 1988) . Независимо от това, псевдокетогенезата може да бъде незначителна в повечето контексти (Bailey et al., 1990; Keller et al., 1978). Схема (фиг. 2А) показва полезен подход, който да се приложи, като се има предвид повишената тъканна концентрация на кетони в стационарно състояние.
Наскоро бъбреците бяха обърнати внимание като потенциално кетогенен орган. В по-голямата част от щатите бъбрекът е нетен консуматор на кетонни тела, извлечени от черния дроб, отделяйки или реабсорбиращи кетонни тела от кръвния поток, а бъбреците обикновено не са генератор или концентратор на нетни кетонни тела (Robinson and Williamson, 1980). Авторите на класическо проучване стигат до заключението, че минималната бъбречна кетогенеза, определена количествено в изкуствена експериментална система, не е физиологично релевантна (Weidemann and Krebs, 1969). Напоследък се прави извод за бъбречна кетогенеза при миши модели с диабет и дефицит на аутофагия, но е по-вероятно многоорганните промени в метаболитната хомеостаза да променят интегративния кетогенен метаболизъм чрез въвеждане на множество органи (Takagi et al., 2016a; Takagi et al., 2016b; Zhang et al., 2011). Една скорошна публикация предлага бъбречната кетогенеза като защитен механизъм срещу исхемично-реперфузионно увреждане в бъбреците (Tran et al., 2016). Абсолютните стационарни концентрации на ?OHB от екстракти от бъбречна тъкан на мишки са докладвани при ~4 mM. За да проверим дали това е приемливо, ние определихме количествено концентрациите на ?OHB в бъбречните екстракти от хранени и 12-часови гладни мишки. Серумните концентрации на ?OHB се увеличават от ~24 µM до 100 mM с 2-часово гладуване (фиг. 24B), докато концентрациите на βOHB в стационарно състояние в бъбреците се приближават до 2 µM в състояние на хранене и само 100 mM при 1-часово гладуване (фиг. 24C E), наблюдения, които са в съответствие с концентрациите, определени количествено преди повече от 2 години (Hems and Brosnan, 45). Остава възможно, че в кетозни състояния, кетонните тела, получени от черния дроб, биха могли да бъдат ренопротективни, но доказателствата за бъбречна кетогенеза изискват допълнително обосноваване. Убедителни доказателства, които подкрепят истинската екстрахепатална кетогенеза, бяха представени в RPE (Adijanto et al., 1970). Предполага се, че тази интригуваща метаболитна трансформация потенциално позволява на кетони, получени от RPE, да текат към фоторецепторните или клетките на глия на Mller, което може да помогне за регенерацията на външния сегмент на фоторецептора.
?OHB като сигнален посредник
Въпреки че са енергийно богати, кетонните тела упражняват провокативна „неканонична” сигнална роля в клетъчната хомеостаза (фиг. 3) (Newman and Verdin, 2014; Rojas-Morales et al., 2016). Например, ?OHB инхибира HDACs от клас I, което увеличава ацетилирането на хистони и по този начин индуцира експресията на гени, които ограничават оксидативния стрес (Shimazu et al., 2013). Самият ?OHB е хистонов ковалентен модификатор на лизинови остатъци в черния дроб на гладни или индуцирани от стрептозотоцин диабетни мишки (Xie et al., 2016) (вижте също по-долу, Интегрирането на метаболизма на кетонните тела, посттранслационна модификация и клетъчна физиология, и Кетонни тела, оксидативен стрес и невропротекция).
�
?OHB също е ефектор чрез G-протеин свързани рецептори. Чрез неясни молекулярни механизми, той потиска активността на симпатиковата нервна система и намалява общия разход на енергия и сърдечната честота чрез инхибиране на сигнализирането на късоверижни мастни киселини чрез G протеин-свързан рецептор 41 (GPR41) (Kimura et al., 2011). Един от най-изследваните сигнални ефекти на ?OHB протича чрез GPR109A (известен също като HCAR2), член на подсемейството на хидрокарбоксилната киселина GPCR, експресиран в мастната тъкан (бяла и кафява) (Tunaru et al., 2003) и в имунни клетки (Ahmed et al., 2009). ?OHB е единственият известен ендогенен лиганд на GPR109A рецептор (EC50 ~770 µM), активиран от d-?OHB, l-?OHB и бутират, но не и AcAc (Taggart et al., 2005). Високият праг на концентрация за активиране на GPR109A се постига чрез придържане към кетогенна диета, гладуване или по време на кетоацидоза, което води до инхибиране на липолизата на мастната тъкан. Антилиполитичният ефект на GPR109A протича чрез инхибиране на аденилциклазата и понижен сАМР, инхибирайки чувствителната към хормона триглицеридна липаза (Ahmed et al., 2009; Tunaru et al., 2003). Това създава отрицателна обратна връзка, в която кетозата поставя модулаторна спирачка на кетогенезата, като намалява освобождаването на неестерифицирани мастни киселини от адипоцитите (Ahmed et al., 2009; Taggart et al., 2005), ефект, който може да бъде уравновесен от симпатикусът, който стимулира липолизата. Ниацинът (витамин В3, никотинова киселина) е мощен (EC50 ~ 0.1 µM) лиганд за GRP109A, ефективно използван от десетилетия за дислипидемии (Benyo et al., 2005; Benyo et al., 2006; Fabbrini et al., 2010; Лукасова и др., 2011; Тунару и др., 2003). Докато ниацинът подобрява обратния транспорт на холестерол в макрофагите и намалява атеросклеротични лезии (Lukasova et al., 2011), ефектите на ?OHB върху атеросклеротични лезии остават неизвестни. Въпреки че GPR109A рецепторът изпълнява защитни роли и съществуват интригуващи връзки между употребата на кетогенна диета при инсулт и невродегенеративни заболявания (Fu et al., 2015; Rahman et al., 2014), защитната роля на ?OHB чрез GPR109A не е демонстрирана in vivo .
И накрая, ?OHB може да повлияе на апетита и ситост. Мета-анализ на проучвания, които измерват ефектите от кетогенните и много нискоенергийни диети, стига до заключението, че участниците, консумиращи тези диети, показват по-висока ситост в сравнение с контролните диети (Gibson et al., 2015). Въпреки това, правдоподобно обяснение за този ефект са допълнителните метаболитни или хормонални елементи, които могат да модулират апетита. Например, мишки, поддържани на кетогенна диета за гризачи, показват повишен разход на енергия в сравнение с мишки, хранени с контролно хранене, въпреки сходния калориен прием и циркулиращият лептин или гените на пептиди, регулиращи поведението на хранене, не са променени (Kennedy et al., 2007). Сред предложените механизми, които предполагат потискане на апетита от ?OHB, включва както сигнализиране, така и окисляване (Laeger et al., 2010). Хепатоцитно специфично заличаване на гена на циркадния ритъм (Per2) и изследванията за имунопреципитация на хроматин разкриват, че PER2 директно активира гена Cpt1a и индиректно регулира Hmgcs2, което води до нарушена кетоза при Per2 нокаут мишки (Chavan et al.,). Тези мишки показаха нарушено очакване на храна, което беше частично възстановено чрез системно приложение на ?OHB. Ще са необходими бъдещи изследвания, за да се потвърди централната нервна система като директна ?OHB мишена и дали е необходимо кетонно окисление за наблюдаваните ефекти или дали е включен друг сигнален механизъм. Други изследователи се позовават на възможността за локална кетогенеза, получена от астроцити във вентромедиалния хипоталамус като регулатор на приема на храна, но тези предварителни наблюдения също ще се възползват от генетични и базирани на потока оценки (Le Foll et al., 2016). Връзката между кетозата и недостига на хранителни вещества остава интерес, тъй като гладът и ситото са важни елементи при неуспешните опити за отслабване.
Интегриране на метаболизма на кетонното тяло, посттранслационна модификация и клетъчна физиология
Кетонните тела допринасят за разделени пулове от ацетил-КоА, ключов междинен продукт, който проявява важна роля в клетъчния метаболизъм (Pietrocola et al., 2015). Една роля на ацетил-CoA е да служи като субстрат за ацетилиране, ензимно катализирана хистонова ковалентна модификация (Choudhary et al., 2014; Dutta et al., 2016; Fan et al., 2015; Menzies et al., 2016 ). Голям брой динамично ацетилирани митохондриални протеини, много от които могат да се появят чрез неензимни механизми, също се появиха от изчислителни протеомични изследвания (Dittenhafer-Reed et al., 2015; Hebert et al., 2013; Rardin et al., 2013 ; Shimazu et al., 2010). Лизин деацетилазите използват цинков кофактор (напр. нуклеоцитозолни HDAC) или NAD+ като ко-субстрат (сиртуини, SIRT) (Choudhary et al., 2014; Menzies et al., 2016). Ацетилпротеомът служи както като сензор, така и като ефектор на общия клетъчен ацетил-КоА пул, тъй като физиологичните и генетичните манипулации водят до неензимни глобални вариации на ацетилирането (Weinert et al., 2014). Тъй като вътреклетъчните метаболити служат като модулатори на ацетилирането на лизинови остатъци, важно е да се вземе предвид ролята на кетонните тела, чието изобилие е силно динамично.
?OHB е епигенетичен модификатор чрез поне два механизма. Повишените нива на ?OHB, предизвикани от гладуване, ограничаване на калориите, директно приложение или продължително упражнение, провокират инхибиране на HDAC или активиране на хистон ацетилтрансфераза (Marosi et al., 2016; Sleiman et al., 2016) или до оксидативен стрес (Shimazu et al.), 2013 3. . Инхибирането на ?OHB на HDAC2016 може да регулира метаболитната физиология на новороденото (Rando et al., 2016). Независимо, самият ?OHB директно модифицира остатъците от хистон лизин (Xie et al., XNUMX). Продължителното гладуване или предизвиканата от стептозотоцин диабетна кетоацидоза повишава хистонова α-хидроксибутирилация. Въпреки че броят на местата на лизин а-хидроксибутирилиране и ацетилиране е сравним, се наблюдава стехиометрично по-голямо хистоново р-хидроксибутирилиране, отколкото ацетилиране. Различни гени са повлияни от хистон лизин а-хидроксибутирилиране, срещу ацетилиране или метилиране, което предполага различни клетъчни функции. Дали а-хидроксибутирилирането е спонтанно или ензимно, не е известно, но разширява обхвата от механизми чрез кетонни тела, които динамично влияят на транскрипцията.
Събитията на съществено клетъчно препрограмиране по време на ограничаване на калориите и липса на хранителни вещества могат да бъдат медиирани в SIRT3- и SIRT5-зависимо митохондриално деацетилиране и десукцинилиране, съответно, регулирайки кетогенните и кетолитичните протеини на пост-транслационно ниво в черния дроб и екстрахепаталните тъкани на ReDittenhafer, 2015; Hebert et al., 2013; Rardin et al., 2013; Shimazu et al., 2010). Въпреки че стехиометричното сравнение на заетите места не е непременно свързано директно с промените в метаболитния поток, митохондриалното ацетилиране е динамично и може да се задвижва от концентрацията на ацетил-КоА или митохондриалното рН, а не от ензимните ацетилтрансферази (Wagner and Payne, 2013). Това, че SIRT3 и SIRT5 модулират активността на ензимите, метаболизиращи кетонното тяло, провокира въпроса за реципрочната роля на кетоните при извайването на ацетилпротеома, сукцинилпротеома и други динамични клетъчни цели. Всъщност, тъй като вариациите на кетогенезата отразяват концентрациите на NAD+, производството и изобилието на кетони могат да регулират активността на сиртуин, като по този начин повлияят на общите пулове ацетил-CoA/сукцинил-CoA, ацилпротеома и по този начин митохондриалната и клетъчната физиология. а-хидроксибутирилирането на остатъци от ензим лизин може да добави още един слой към клетъчното препрограмиране. В екстрахепаталните тъкани окисляването на кетонните тела може да стимулира аналогични промени в клетъчната хомеостаза. Докато компартментацията на пуловете на ацетил-CoA е силно регулирана и координира широк спектър от клетъчни промени, способността на кетонните тела да оформят директно концентрациите на ацетил-CoA в митохондриите и цитоплазмата изисква изясняване (Chen et al., 2012; Corbet et al., 2016; Pougovkina et al., 2014; Schwer et al., 2009; Wellen and Thompson, 2012). Тъй като концентрациите на ацетил-КоА са строго регулирани, а ацетил-КоА е непроницаем за мембраната, от решаващо значение е да се вземат предвид двигателните механизми, координиращи хомеостазата на ацетил-КоА, включително скоростта на производство и терминално окисление в TCA цикъла, превръщане в кетонни тела, митохондриални изтичане чрез карнитин ацетилтрансфераза (CrAT) или износ на ацетил-КоА в цитозола след превръщане в цитрат и освобождаване от ATP цитрат лиаза (ACLY). Ключовите роли на тези последни механизми в клетъчния ацетилпротеом и хомеостазата изискват съвпадащо разбиране на ролите на кетогенезата и окислението на кетоните (Das et al., 2015; McDonnell et al., 2016; Moussaieff et al., 2015; Overmyer et al., 2015; Seiler et al., 2014; Seiler et al., 2015; Wellen et al., 2009; Wellen и Thompson, 2012). Ще са необходими конвергентни технологии в метаболомиката и ацилпротеомиката в условията на генетично манипулирани модели, за да се определят цели и резултати.
Противовъзпалителни и противовъзпалителни реакции към кетонните тела
Кетозата и кетонните тела модулират възпалението и функцията на имунните клетки, но са предложени различни и дори несъответстващи механизми. Продължителното недостиг на хранителни вещества намалява възпалението (Youm et al., 2015), но хроничната кетоза на диабет тип 1 е провъзпалително състояние (Jain et al., 2002; Kanikarla-Marie and Jain, 2015; Kurepa et al., 2012 ). Основаните на механизма сигнални роли за ?OHB при възпаление се появяват, защото много клетки на имунната система, включително макрофаги или моноцити, обилно експресират GPR109A. Докато ?OHB упражнява предимно противовъзпалителен отговор (Fu et al., 2014; Gambhir et al., 2012; Rahman et al., 2014; Youm et al., 2015), високи концентрации на кетонни тела, особено AcAc, могат предизвикват провъзпалителен отговор (Jain et al., 2002; Kanikarla-Marie and Jain, 2015; Kurepa et al., 2012).
Противовъзпалителните роли на GPR109A лигандите при атеросклероза, затлъстяване, възпалително заболяване на червата, неврологични заболявания и рак са разгледани (Graff et al., 2016). Експресията на GPR109A се увеличава в RPE клетки на диабетни модели, пациенти с диабет при хора (Gambhir et al., 2012) и в микроглия по време на невродегенерация (Fu et al., 2014). Противовъзпалителните ефекти на ?OHB се засилват от свръхекспресията на GPR109A в клетките на RPE и се отменят чрез фармакологично инхибиране или генетично нокаутиране на GPR109A (Gambhir et al., 2012). ?OHB и екзогенна никотинова киселина (Taggart et al., 2005), и двете придават противовъзпалителни ефекти в TNF? или LPS-индуцирано възпаление чрез намаляване на нивата на провъзпалителни протеини (iNOS, COX-2) или секретирани цитокини (TNFa, IL-1a, IL-6, CCL2/MCP-1), отчасти чрез инхибиране на NF -?B транслокация (Fu et al., 2014; Gambhir et al., 2012). ?OHB намалява ER стреса и NLRP3 инфламазомата, активирайки реакцията на антиоксидантен стрес (Bae et al., 2016; Youm et al., 2015). Въпреки това, при невродегенеративно възпаление, GPR109A-зависимата ?OHB-медиирана защита не включва възпалителни медиатори като сигнализиране на пътя на MAPK (напр. ERK, JNK, p38) (Fu et al., 2014), но може да изисква COX-1-зависим PGD2 производство (Rahman et al., 2014). Интригуващо е, че макрофагът GPR109A е необходим за упражняване на невропротективен ефект в модел на исхемичен инсулт (Rahman et al., 2014), но способността на ?OHB да инхибира възпалението на NLRP3 в макрофагите, получени от костния мозък, е независима от GPR109A ., 2015 г.). Въпреки че повечето проучвания свързват ?OHB с противовъзпалителни ефекти, ?OHB може да е провъзпалително и да повишава маркерите на липидна пероксидация в хепатоцитите на телетата (Shi et al., 2014). По този начин противовъзпалителният ефект на αOHB може да зависи от типа на клетката, концентрацията на αOHB, продължителността на експозиция и наличието или отсъствието на ко-модулатори.
За разлика от ?OHB, AcAc може да активира про-възпалителна сигнализация. Повишеният AcAc, особено с висока концентрация на глюкоза, засилва увреждането на ендотелните клетки чрез механизъм, зависим от NADPH оксидаза/оксидативен стрес (Kanikarla-Marie and Jain, 2015). Високите концентрации на AcAc в пъпната връв на майки с диабет са свързани с по-висока скорост на окисление на протеини и концентрация на MCP-1 (Kurepa et al., 2012). Високият AcAc при пациенти с диабет е свързан с TNF? експресия (Jain et al., 2002) и AcAc, но не ?OHB, индуцира TNF?, MCP-1 експресия, натрупване на ROS и намалено ниво на cAMP в U937 човешки моноцитни клетки (Jain et al., 2002; Kurepa et al. ., 2012 г.).
Сигналните явления, зависими от кетонното тяло, често се задействат само при високи концентрации на кетонно тяло (> 5 mM) и в случай на много проучвания, свързващи кетоните с про- или противовъзпалителни ефекти, чрез неясни механизми. В допълнение, поради противоречивите ефекти на ?OHB срещу AcAc върху възпалението и способността на съотношението AcAc/?OHB да повлияе на митохондриалния редокс потенциал, най-добрите експерименти, оценяващи ролите на кетонните тела върху клетъчните фенотипове, сравняват ефектите на AcAc и ? OHB в различни съотношения и при различни кумулативни концентрации [напр. (Saito et al., 2016)]. И накрая, AcAc може да бъде закупен в търговската мрежа само като литиева сол или като етилов естер, който изисква основна хидролиза преди употреба. Литиевият катион независимо индуцира каскади на сигнална трансдукция (Manji et al., 1995), а AcAc анионът е лабилен. И накрая, изследванията, използващи рацемичен d/l-?OHB, могат да бъдат объркани, тъй като само d-?OHB стереоизомерът може да бъде окислен до AcAc, но d-?OHB и l-?OHB могат всеки да сигнализира чрез GPR109A, да инхибира NLRP3 инфламазомата, и служат като липогенни субстрати.
Кетонни тела, оксидативен стрес и неврозащита
Оксидативният стрес обикновено се дефинира като състояние, в което ROS са представени в излишък, поради прекомерно производство и/или нарушено елиминиране. Ролите на кетонните тела за смекчаване на антиоксидантния и оксидативния стрес са широко описани както in vitro, така и in vivo, особено в контекста на невропротекцията. Тъй като повечето неврони не генерират ефективно високоенергийни фосфати от мастни киселини, но окисляват кетонните тела, когато въглехидратите са в недостиг, невропротективните ефекти на кетонните тела са особено важни (Cahill GF Jr, 2006; Edmond et al., 1987; Yang et al., 1987). В моделите на оксидативен стрес индукцията на BDH1 и потискането на SCOT предполагат, че метаболизмът на кетоновите тела може да бъде препрограмиран, за да поддържа разнообразна клетъчна сигнализация, редокс потенциал или метаболитни изисквания (Nagao et al., 2016; Tieu et al., 2003).
Кетонните тела намаляват степента на клетъчно увреждане, нараняване, смърт и по-ниска апоптоза в неврони и кардиомиоцити (Haces et al., 2008; Maalouf et al., 2007; Nagao et al., 2016; Tieu et al., 2003). Извикваните механизми са разнообразни и не винаги са линейно свързани с концентрацията. Ниските милимоларни концентрации на (d или l)-?OHB улавят ROS (хидроксил анион), докато AcAc улавя множество видове ROS, но само при концентрации, които надхвърлят физиологичния диапазон (IC50 20 mM) (Haces et al., 67) . Обратно, благоприятното влияние върху окислително-редукционния потенциал на веригата за транспортиране на електрони е механизъм, обикновено свързан с d-?OHB. Докато и трите кетонни тела (d/l-?OHB и AcAc) намаляват смъртта на невронните клетки и натрупването на ROS, предизвикано от химическо инхибиране на гликолизата, само d-?OHB и AcAc предотвратяват намаляването на невроналния АТФ. Обратно, в хипогликемичен in vivo модел, (d или l)-?OHB, но не и AcAc предотвратяват хипокампалната липидна пероксидация (Haces et al., 2008; Maalouf et al., 2008; Marosi et al., 2007; Murphy, 2016 ; Tieu et al., 2009). Изследвания in vivo на мишки, хранени с кетогенна диета (2003% kcal мазнини и 87% протеин) показват невроанатомични вариации на антиоксидантния капацитет (Ziegler et al., 13), където най-дълбоките промени се наблюдават в хипокампуса, с повишаване на глутатион пероксидазата и общата антиоксидантни способности.
Кетогенната диета, кетонните естери (вижте също Терапевтично използване на кетогенна диета и екзогенни кетонни тела) или прилагането на ?OHB оказват невропротекция при модели на исхемичен инсулт (Rahman et al., 2014); болест на Паркинсон (Tieu et al., 2003); припадък на кислородна токсичност на централната нервна система (D'Agostino et al., 2013); епилептични спазми (Yum et al., 2015); синдром на митохондриална енцефаломиопатия, лактатна ацидоза и подобни на инсулт (MELAS) епизоди (Frey et al., 2016) и болест на Алцхаймер (Cunnane and Crawford, 2003; Yin et al., 2016). Обратно, скорошен доклад демонстрира хистопатологични доказателства за невродегенеративна прогресия чрез кетогенна диета в модел на трансгенна мишка на анормално възстановяване на митохондриална ДНК, въпреки увеличаването на митохондриалната биогенеза и антиоксидантните сигнатури (Lauritzen et al., 2016). Други противоречиви доклади предполагат, че излагането на високи концентрации на кетонни тела предизвиква оксидативен стрес. Високите дози ?OHB или AcAc индуцират секреция на азотен оксид, липидна пероксидация, намалена експресия на SOD, глутатион пероксидаза и каталаза в хепатоцитите на телета, докато в хепатоцитите на плъхове индукцията на MAPK пътя се приписва на AcAc, но не и на ?OHB (Abdelmegeed et al. ; Shi et al., 2004; Shi et al., 2014).
Взети заедно, повечето доклади свързват ?OHB с отслабването на оксидативния стрес, тъй като приложението му инхибира производството на ROS/супероксид, предотвратява липидна пероксидация и окисляване на протеини, повишава нивата на антиоксидантния протеин и подобрява митохондриалното дишане и производството на АТФ (Abdelmegeed et al., 2004); Haces et al., 2008; Jain et al., 1998; Jain et al., 2002; Kanikarla-Marie and Jain, 2015; Maalouf et al., 2007; Maalouf и Rho, 2008; Marosi et al., 2016; Tieu et al., 2003; Yin et al., 2016; Ziegler et al., 2003). Докато AcAc е по-пряко свързан от ?OHB с индуцирането на оксидативен стрес, тези ефекти не винаги се разграничават лесно от проспективните провъзпалителни отговори (Jain et al., 2002; Kanikarla-Marie and Jain, 2015; Kanikarla-Marie и Джайн, 2016 г.). Освен това е от решаващо значение да се има предвид, че очевидната антиоксидантна полза, предоставена от плейотропните кетогенни диети, може да не бъде трансдуцирана от самите кетонни тела, а невропротекцията, предоставена от кетонните тела, може да не се дължи изцяло на оксидативния стрес. Например по време на лишаване от глюкоза, в модел на лишаване от глюкоза в кортикалните неврони, ?OHB стимулира автофагичния поток и предотвратява натрупването на автофагозома, което е свързано с намалена невронна смърт (Camberos-Luna et al., 2016). d-?OHB индуцира също и каноничните антиоксидантни протеини FOXO3a, SOD, MnSOD и каталаза, проспективно чрез HDAC инхибиране (Nagao et al., 2016; Shimazu et al., 2013).
Безалкохолна мастна чернодробна болест (NAFLD) и метаболизъм на кетонното тяло
Свързаната със затлъстяването NAFLD и неалкохолен стеатохепатит (NASH) са най-честите причини за чернодробно заболяване в западните страни (Rinella and Sanyal, 2016), а индуцираната от NASH чернодробна недостатъчност е една от най-честите причини за чернодробна трансплантация. Докато излишното съхранение на триацилглицероли в хепатоцити >5% от теглото на черния дроб (NAFL) само по себе си не причинява дегенеративна чернодробна функция, прогресията към NAFLD при хората корелира със системна инсулинова резистентност и повишен риск от диабет тип 2 и може да допринесе за патогенезата на сърдечно-съдови заболявания и хронично бъбречно заболяване (Fabbrini et al., 2009; Targher et al., 2010; Targher and Byrne, 2013). Патогенните механизми на NAFLD и NASH не са напълно разбрани, но включват аномалии на хепатоцитния метаболизъм, хепатоцитната аутофагия и стрес на ендоплазмения ретикулум, функцията на чернодробните имунни клетки, възпаление на мастната тъкан и системни възпалителни медиатори (Fabbrini et al., 2009, Masaniu, 2013; ; Targher et al., 2010; Yang et al., 2010). Смущенията в метаболизма на въглехидратите, липидите и аминокиселините се появяват и допринасят за затлъстяване, диабет и NAFLD при хора и в моделни организми [прегледани в (Farese et al., 2012; Lin and Accili, 2011; Newgard, 2012; Samuel и Шулман, 2012; Слънце и Лазар, 2013)]. Докато хепатоцитните аномалии в метаболизма на цитоплазмените липиди често се наблюдават при NAFLD (Fabbrini et al., 2010b), ролята на митохондриалния метаболизъм, който управлява окислителното изхвърляне на мазнините, е по-малко ясна в патогенезата на NAFLD. Аномалии на митохондриалния метаболизъм се появяват и допринасят за патогенезата на NAFLD/NASH (Hyotylainen et al., 2016; Serviddio et al., 2011; Serviddio et al., 2008; Wei et al., 2008). Има общо (Felig et al., 1974; Iozzo et al., 2010; Koliaki et al., 2015; Satapati et al., 2015; Satapati et al., 2012; Sunny et al., 2011), но не е еднородно ( Koliaki и Roden, 2013; Perry et al., 2016; Rector et al., 2010) консенсус, че преди развитието на добросъвестния NASH, чернодробното митохондриално окисление и по-специално окисляването на мазнините се увеличава при затлъстяване, системна инсулинова резистентност и NAFLD. Вероятно е, че с напредването на NAFLD се появява хетерогенност на окислителния капацитет, дори сред отделните митохондрии, и в крайна сметка оксидативната функция се нарушава (Koliaki et al., 2015; Rector et al., 2010; Satapati et al., 2008; Satapati et al. ., 2012 г.).
Кетогенезата често се използва като заместител на окисляването на чернодробните мазнини. Нарушенията на кетогенезата се появяват с напредването на NAFLD при животински модели и вероятно при хора. Чрез непълно дефинирани механизми, хиперинсулинемията потиска кетогенезата, вероятно допринасяйки за хипокетонемия в сравнение с слабите контроли (Bergman et al., 2007; Bickerton et al., 2008; Satapati et al., 2012; Soeters2009 et al., , ; , 2011; Vice et al., 2005). Независимо от това, способността на концентрациите на циркулиращи кетонни тела да предскажат NAFLD е противоречива (M nnist et al., 2015; Sanyal et al., 2001). Стабилните количествени магнитно-резонансни спектроскопски методи в животински модели разкриват повишена скорост на обмяна на кетони с умерена инсулинова резистентност, но намалените нива са очевидни при по-тежка инсулинова резистентност (Satapati et al., 2012; Sunny et al., 2010). При затлъстели хора с мастен черен дроб кетогенната скорост е нормална (Bickerton et al., 2008; Sunny et al., 2011) и следователно скоростта на кетогенезата е намалена спрямо увеличеното натоварване на мастните киселини в хепатоцитите. Следователно, ацетил-КоА, извлечен от α-окисление, може да бъде насочен към терминално окисление в TCA цикъла, увеличавайки терминалното окисление, глюконеогенеза, задвижвана от фосфоенолпируват чрез анаплероза/катаплероза и оксидативен стрес. Ацетил-КоА също вероятно претърпява износ от митохондриите като цитрат, прекурсорен субстрат за липогенезата (фиг. 4) (Satapati et al., 2015; Satapati et al., 2012; Solinas et al., 2015). Докато кетогенезата става по-малко чувствителна към инсулин или гладуване с продължително затлъстяване (Satapati et al., 2012), основните механизми и последиците от това надолу по веригата остават ненапълно разбрани. Последните данни показват, че mTORC1 потиска кетогенезата по начин, който може да бъде след инсулиновата сигнализация (Kucejova et al., 2016), което е в съответствие с наблюденията, че mTORC1 инхибира PPAR?-медиирана индукция на Hmgcs2 (Sengupta et al., 2010). също вижте Регламент на HMGCS2 и SCOT/OXCT1).
�
Предварителните наблюдения от нашата група предполагат неблагоприятни чернодробни последици от кетогенната недостатъчност (Cotter et al., 2014). За да проверим хипотезата, че нарушената кетогенеза, дори в състояния, наситени с въглехидрати и по този начин „некетогенни“ състояния, допринася за анормален метаболизъм на глюкозата и провокира стеатохепатит, ние генерирахме миши модел на изразена кетогенна недостатъчност чрез прилагане на антисенс олигонукле (ASO) Hmgcs2. Загубата на HMGCS2 при възрастни мишки, хранени с ниско съдържание на мазнини, причинява лека хипергликемия и значително повишено производство на стотици чернодробни метаболити, набор от които силно предполага активиране на липогенезата. Диетично хранене с високо съдържание на мазнини на мишки с недостатъчна кетогенеза води до обширно увреждане и възпаление на хепатоцитите. Тези констатации подкрепят централните хипотези, че (i) кетогенезата не е пасивен път на преливане, а по-скоро динамичен възел в чернодробната и интегрирана физиологична хомеостаза, и (ii) разумното кетогенно увеличаване за смекчаване на NAFLD/NASH и нарушен метаболизъм на чернодробната глюкоза е достойно за изследване .
Как може нарушената кетогенеза да допринесе за чернодробно увреждане и променена глюкозна хомеостаза? Първото съображение е дали виновникът е дефицитът на кетогенен поток или самите кетони. Неотдавнашен доклад предполага, че кетонните тела могат да смекчат индуцираното от оксидативен стрес чернодробно увреждане в отговор на n-3 полиненаситени мастни киселини (Pawlak et al., 2015). Припомнете си, че поради липса на експресия на SCOT в хепатоцитите, кетонните тела не се окисляват, но те могат да допринесат за липогенезата и да служат на различни сигнални роли, независимо от тяхното окисление (вижте също неокислителни метаболитни съдби на кетонни тела и ?OHB като сигнален медиатор). Възможно е също така получените от хепатоцити кетонни тела да служат като сигнал и/или метаболит за съседни типове клетки в чернодробния ацинус, включително звездни клетки и макрофаги на Купферова клетка. Докато ограничената налична литература предполага, че макрофагите не са в състояние да окисляват кетонни тела, това е измерено само с помощта на класически методологии и само в перитонеални макрофаги (Newsholme et al., 1986; Newsholme et al., 1987), което показва, че повторното оценката е подходяща, като се има предвид изобилната експресия на SCOT в макрофаги, получени от костен мозък (Youm et al., 2015).
Хепатоцитният кетогенен поток може също да бъде цитопротективен. Докато оздравителните механизми може да не зависят от кетогенезата сама по себе си, кетогенните диети с ниско съдържание на въглехидрати са свързани с подобряване на NAFLD (Browning et al., 2011; Foster et al., 2010; Kani et al., 2014; Schugar and Crawford, 2012) . Нашите наблюдения показват, че кетогенезата на хепатоцитите може да даде обратна връзка и да регулира потока на TCA цикъла, анаплеротичния поток, глюконеогенезата, получена от фосфоенолпируват (Cotter et al., 2014) и дори оборота на гликоген. Кетогенното увреждане насочва ацетил-КоА към увеличаване на потока на TCA, който в черния дроб е свързан с повишено ROS-медиирано увреждане (Satapati et al., 2015; Satapati et al., 2012); принуждава отклоняване на въглерод в de novo синтезирани липидни видове, които могат да се окажат цитотоксични; и предотвратява повторното окисление на NADH до NAD+ (Cotter et al., 2014) (фиг. 4). Взети заедно, бъдещи експерименти са необходими за справяне с механизмите, чрез които относителната кетогенна недостатъчност може да стане неадаптивна, да допринесе за хипергликемия, да провокира стеатохепатит и дали тези механизми функционират при човешки NAFLD/NASH. Тъй като епидемиологичните данни предполагат нарушена кетогенеза по време на прогресията на стеатохепатит (Embade et al., 2016; Marinou et al., 2011; M nnist et al., 2015; Pramfalk et al., 2015; Safaei et al., 2016) терапиите, които повишават чернодробната кетогенеза, могат да се окажат полезни (Degirolamo et al., 2016; Honda et al., 2016).
Кетонни тела и сърдечна недостатъчност (HF)
С метаболитна скорост над 400 kcal/kg/ден и оборот от 6 kg ATP/ден, сърцето е органът с най-висок разход на енергия и окислително търсене (Ashrafian et al., 35; Wang et al., 2007б). По-голямата част от енергийния оборот на миокарда се намира в митохондриите, а 2010% от това снабдяване произхожда от ФАО. Сърцето е всеядно и гъвкаво при нормални условия, но патологично ремоделиращото се сърце (например поради хипертония или инфаркт на миокарда) и диабетното сърце стават метаболитно негъвкави (Balasse and Fery, 70; BING, 1989; Fukao et al., 1954 ; Lopaschuk et al., 2004; Taegtmeyer et al., 2010; Taegtmeyer et al., 1980; Young et al., 2002). Всъщност генетично програмираните аномалии на сърдечния горивен метаболизъм при миши модели провокират кардиомиопатия (Carley et al., 2002; Neubauer, 2014). При физиологични условия нормалните сърца окисляват кетонните тела пропорционално на доставянето им, за сметка на окисляването на мастни киселини и глюкоза, а миокардът е най-големият консуматор на кетонно тяло на единица маса (BING, 2007; Crawford et al., 1954; GARLAND et al. ., 2009; Hasselbaink et al., 1962; Jeffrey et al., 2003; Pelletier et al., 1995; Tardif et al., 2007; Yan et al., 2001). В сравнение с окисляването на мастни киселини, кетонните тела са по-енергийно ефективни, давайки повече енергия, налична за синтеза на АТФ на молекула вложен кислород (P/O съотношение) (Kashiwaya et al., 2009; Sato et al., 2010; Veech, 1995) . Окислението на кетонните тела също води до потенциално по-висока енергия от FAO, поддържайки окисление на убихинона, което увеличава редокс обхвата във веригата за транспорт на електрони и прави повече енергия налична за синтезиране на ATP (Sato et al., 2004; Veech, 1995). Окислението на кетонните тела може също да намали производството на ROS и по този начин оксидативния стрес (Veech, 2004).
Предварителните интервенционни и наблюдателни проучвания показват потенциална оздравителна роля на кетонните тела в сърцето. В контекста на експериментална исхемия/реперфузионно увреждане, кетонните тела придават потенциални кардиопротективни ефекти (Al-Zaid et al., 2007; Wang et al., 2008), вероятно поради увеличаването на изобилието на митохондриите в сърцето или повишаването на регулирането на решаващо окислително фосфорилиране медиатори (Snorek et al., 2012; Zou et al., 2002). Последните проучвания показват, че използването на кетонни тела се увеличава при неуспешни сърца на мишки (Aubert et al., 2016) и хора (Bedi et al., 2016), подкрепяйки предишни наблюдения при хора (BING, 1954; Fukao et al., 2000; Janardhan et al., 2011; Longo et al., 2004; Rudolph and Schinz, 1973; Tildon и Cornblath, 1972). Концентрациите на циркулиращите кетонни тела се повишават при пациенти със сърдечна недостатъчност, право пропорционално на налягането на пълнене, наблюдения, чийто механизъм и значение остават неизвестни (Kupari et al., 1995; Lommi et al., 1996; Lommi et al., 1997; Neely et al. ., 1972), но мишките със селективен дефицит на SCOT в кардиомиоцитите показват ускорено патологично камерно ремоделиране и ROS сигнатури в отговор на хирургично индуцирано увреждане от претоварване с налягане (Schugar et al., 2014).
Последните интригуващи наблюдения в терапията на диабета разкриха потенциална връзка между метаболизма на миокарда на кетони и патологичното камерно ремоделиране (Фиг. 5). Инхибирането на бъбречния проксимален тубулен натриев/глюкозен ко-транспортер 2 (SGLT2i) повишава концентрациите на циркулиращи кетонни тела при хора (Ferrannini et al., 2016a; Inagaki et al., 2015) и мишки (Suzuki et al., 2014) чернодробна кетогенеза (Ferrannini et al., 2014; Ferrannini et al., 2016a; Katz и Leiter, 2015; Mudaliar et al., 2015). Поразително е, че поне един от тези агенти намалява хоспитализацията при сърдечна недостатъчност (напр., както е разкрито от проучването EMPA-REG OUTOME) и подобрява сърдечно-съдовата смъртност (Fitchett et al., 2016; Sonesson et al., 2016; Wu et al., 2016a ; Zinman et al., 2015). Докато движещите механизми зад благоприятните резултати от СН за свързания SGLT2i остават активно обсъждани, ползата от оцеляването вероятно е многофакторна, проспективно включваща кетоза, но също и благотворни ефекти върху теглото, кръвното налягане, нивата на глюкоза и пикочна киселина, артериалната скованост, симпатиковата нервна система, осмотичното диуреза/намален плазмен обем и повишен хематокрит (Raz and Cahn, 2016; Vallon and Thomson, 2016). Взети заедно, схващането, че терапевтично увеличаващата се кетонемия при пациенти със СН или при тези с висок риск от развитие на СН, остава противоречива, но е в процес на активно изследване в предклинични и клинични проучвания (Ferrannini et al., 2016b; Kolwicz et al., 2016; Lopaschuk и Verma, 2016; Mudaliar et al., 2016; Taegtmeyer, 2016).
�
Кетонни тела в биологията на рака
Връзките между кетонните тела и рака бързо се появяват, но проучванията както върху животински модели, така и при хора са довели до различни заключения. Тъй като кетонният метаболизъм е динамичен и реагира на хранителните състояния, примамливо е да се търсят биологични връзки с рака поради потенциала за прецизно ръководени хранителни терапии. Раковите клетки се подлагат на метаболитно препрограмиране, за да поддържат бърза клетъчна пролиферация и растеж (DeNicola and Cantley, 2015; Pavlova and Thompson, 2016). Класическият ефект на Варбург в метаболизма на раковите клетки възниква от доминиращата роля на гликолизата и ферментацията на млечна киселина за пренос на енергия и компенсиране на по-ниската зависимост от окислителното фосфорилиране и ограниченото митохондриално дишане (De Feyter et al., 2016; Grabacka et al., 2016; Kang et al., 2015; Poff et al., 2014; Shukla et al., 2014). Глюкозният въглерод се насочва основно чрез гликолиза, пентозофосфатен път и липогенеза, които заедно осигуряват междинни продукти, необходими за разширяване на туморната биомаса (Grabbacka et al., 2016; Shukla et al., 2014; Yoshii et al., 2015). Адаптирането на раковите клетки към лишаване от глюкоза се осъществява чрез способността да се използват алтернативни източници на гориво, включително ацетат, глутамин и аспартат (Jaworski et al., 2016; Sullivan et al., 2015). Например, ограничен достъп до пируват разкрива способността на раковите клетки да превръщат глутамин в ацетил-КоА чрез карбоксилиране, поддържайки както енергийните, така и анаболните нужди (Yang et al., 2014). Интересна адаптация на раковите клетки е използването на ацетат като гориво (Comerford et al., 2014; Jaworski et al., 2016; Mashimo et al., 2014; Wright and Simone, 2016; Yoshii et al., 2015). Ацетатът също е субстрат за липогенезата, която е от решаващо значение за пролиферацията на туморни клетки, а усилването на този липогенен канал е свързано с по-кратка преживяемост на пациентите и по-голямо туморно натоварване (Comerford et al., 2014; Mashimo et al., 2014; Yoshii et al. ., 2015 г.).
Нераковите клетки лесно прехвърлят своя енергиен източник от глюкоза към кетонни тела по време на лишаване от глюкоза. Тази пластичност може да е по-променлива при видовете ракови клетки, но in vivo имплантирани мозъчни тумори окисляват [2,4-13C2]-?OHB до подобна степен като заобикалящата мозъчна тъкан (De Feyter et al., 2016). Моделите на „обратен ефект на Варбург“ или „туморен метаболизъм с две отделения“ предполагат, че раковите клетки индуцират производството на ?OHB в съседни фибробласти, осигурявайки енергийните нужди на туморните клетки (Bonuccelli et al., 2010; Martinez-Outschoorn et al., 2012) . В черния дроб изместването на хепатоцитите от кетогенеза към окисление на кетони в клетките на хепатоцелуларен карцином (хепатом) е в съответствие с активирането на BDH1 и SCOT активностите, наблюдавани в две хепатомни клетъчни линии (Zhang et al., 1989). Наистина, хепатомните клетки експресират OXCT1 и BDH1 и окисляват кетони, но само когато серумът е гладен (Huang et al., 2016). Като алтернатива е предложена и кетогенеза на туморни клетки. Динамичните промени в експресията на кетогенния ген се проявяват по време на ракова трансформация на епитела на дебелото черво, клетъчен тип, който нормално експресира HMGCS2, а скорошен доклад предполага, че HMGCS2 може да бъде прогностичен маркер за лоша прогноза при колоректални и плоскоклетъчни карциноми (Camarero et al. 2006; Чен и др., 2016). Остава да се определи дали тази асоциация изисква или включва кетогенеза, или съвместна функция на HMGCS2. Обратно, очевидното производство на ?OHB от клетките на меланома и глиобластома, стимулирано от PPAR? агонист фенофибрат, се свързва със спиране на растежа (Grabacka et al., 2016). Необходими са по-нататъшни проучвания, за да се характеризират ролите на експресията на HMGCS2/SCOT, кетогенезата и окисляването на кетони в раковите клетки.
Отвъд сферата на горивния метаболизъм, кетоните наскоро бяха замесени в биологията на раковите клетки чрез сигнален механизъм. Анализът на BRAF-V600E+ меланом показва OCT1-зависима индукция на HMGCL по онкогенен BRAF-зависим начин (Kang et al., 2015). Увеличаването на HMGCL е свързано с по-висока клетъчна концентрация на AcAc, което от своя страна засилва взаимодействието на BRAFV600E-MEK1, усилвайки MEK-ERK сигнализирането в цикъл за предаване, който задвижва пролиферацията и растежа на туморните клетки. Тези наблюдения повдигат интригуващия въпрос за проспективната екстрахепатална кетогенеза, която след това поддържа сигнален механизъм (вижте също ?OHB като сигнален медиатор и Противоречия в екстрахепаталната кетогенеза). Също така е важно да се вземат предвид независимите ефекти на AcAc, d-?OHB и l-?OHB върху метаболизма на рака, а когато се разглежда HMGCL, катаболизмът на левцина също може да бъде нарушен.
Ефектите от кетогенните диети (вижте също Терапевтично използване на кетогенна диета и екзогенни кетонни тела) при ракови животински модели са разнообразни (De Feyter et al., 2016; Klement et al., 2016; Meidenbauer et al., 2015; Poff et al. ., 2014; Seyfried et al., 2011; Shukla et al., 2014). Докато епидемиологичните асоциации между затлъстяването, рака и кетогенните диети се обсъждат (Liskiewicz et al., 2016; Wright and Simone, 2016), мета-анализ, използващ кетогенни диети в животински модели и в проучвания при хора, предполага благотворно въздействие върху оцеляването, с ползи, проспективно свързани с големината на кетозата, времето на започване на диетата и местоположението на тумора (Klement et al., 2016; Woolf et al., 2016). Лечението на ракови клетки на панкреаса с кетонни тела (d-?OHB или AcAc) инхибира растежа, пролиферацията и гликолизата, а кетогенната диета (81% kcal мазнини, 18% протеин, 1% въглехидрати) намалява in vivo теглото на тумора, гликемията и повишено мускулно и телесно тегло при животни с имплантиран рак (Shukla et al., 2014). Подобни резултати са наблюдавани при използване на метастатичен клетъчен модел на глиобластом при мишки, които получават кетонни добавки в диетата (Poff et al., 2014). Обратно, кетогенната диета (91% kcal мазнини, 9% протеин) повишава циркулиращата концентрация на ?OHB и намалява гликемията, но не оказва влияние нито върху обема на тумора, нито върху продължителността на преживяемостта при плъхове, носещи глиома (De Feyter et al., 2016). Предложен е глюкозен кетонов индекс като клиничен индикатор, който подобрява метаболитното управление на терапията на мозъчен рак, предизвикана от кетогенна диета при хора и мишки (Meidenbauer et al., 2015). Взети заедно, ролите на метаболизма на кетонните тела и кетонните тела в биологията на рака са примамливи, защото всеки от тях представлява поносими терапевтични възможности, но основните аспекти остават да бъдат изяснени, като ясни влияния възникват от матрица от променливи, включително (i) разлики между екзогенния кетон тела срещу кетогенна диета, (ii) тип ракови клетки, геномни полиморфизми, степен и стадий; и (iii) времето и продължителността на излагане на кетотично състояние.
Кетогенезата се създава от кетонни тела чрез разграждане на мастни киселини и кетогенни аминокиселини. Този биохимичен процес осигурява енергия на различни органи, по-специално мозъка, при условия на гладуване като отговор на липсата на кръвна глюкоза. Кетонните тела се произвеждат главно в митохондриите на чернодробните клетки. Докато други клетки са способни да извършват кетогенеза, те не са толкова ефективни в това, колкото чернодробните клетки. Тъй като кетогенезата се осъществява в митохондриите, нейните процеси се регулират независимо. Д-р Алекс Хименес DC, CCST Insight
Терапевтично приложение на кетогенна диета и екзогенни кетонни тела
Приложенията на кетогенните диети и кетонните тела като терапевтични инструменти също са възникнали в неракови контексти, включително затлъстяване и NAFLD/NASH (Browning et al., 2011; Foster et al., 2010; Schugar and Crawford, 2012); сърдечна недостатъчност (Huynh, 2016; Kolwicz et al., 2016; Taegtmeyer, 2016); неврологични и невродегенеративни заболявания (Martin et al., 2016; McNally and Hartman, 2012; Rho, 2015; Rogawski et al., 2016; Yang and Cheng, 2010; Yao et al., 2011); вродени нарушения на метаболизма (Scholl-B rgi et al, 2015); и изпълнение на упражненията (Cox et al., 2016). Ефикасността на кетогенните диети е особено оценена при терапията на епилептичен припадък, особено при резистентни към лекарства пациенти. Повечето проучвания оценяват кетогенните диети при педиатрични пациенти и разкриват до ~50% намаление на честотата на припадъци след 3 месеца, с подобрена ефективност при избрани синдроми (Wu et al., 2016b). Опитът е по-ограничен при епилепсия при възрастни, но подобно намаление е очевидно, с по-добър отговор при пациенти с симптоматична генерализирана епилепсия (Nei et al., 2014). Основните антиконвулсивни механизми остават неясни, въпреки че постулираните хипотези включват намалено използване на глюкоза/гликолиза, препрограмиран транспорт на глутамат, индиректно въздействие върху ATP-чувствителния калиев канал или аденозин A1 рецептор, промяна в експресията на изоформата на натриевия канал или ефекти върху циркулиращите хормони (включително лептин). Lambrechts et al., 2016; Lin et al., 2017; Lutas and Yellen, 2013). Остава неясно дали антиконвулсивният ефект се дължи основно на кетонните тела или на каскадните метаболитни последици от диетите с ниско съдържание на въглехидрати. Независимо от това, кетонните естери (вижте по-долу) изглежда повишават прага на гърчове при животински модели на провокирани припадъци (Ciarlone et al., 2016; D'Agostino et al., 2013; Viggiano et al., 2015).
Диети в стил Аткинс и кетогенни, ниско въглехидратни диети често се считат за неприятни и могат да причинят запек, хиперурикемия, хипокалцемия, хипомагнезиемия, да доведат до нефролитиаза, кетоацидоза, да причинят хипергликемия и да повишат концентрацията на циркулиращия холестерол и свободни мастни киселини, ; Kossoff and Hartman, 2001; Kwiterovich et al., 2012; Suzuki et al., 2003). Поради тези причини дългосрочното придържане представлява предизвикателства. Изследванията при гризачи обикновено използват отличително разпределение на макронутриентите (2002% kcal мазнини, 94% kcal въглехидрати, 1% kcal протеин, Bio-Serv F5), което провокира силна кетоза. Въпреки това, увеличаването на съдържанието на протеин, дори до 3666% kcal, значително намалява кетозата, а ограничаването на протеина от 10% kcal придава объркващи метаболитни и физиологични ефекти. Тази диетична формулировка също е изчерпана с холин, друга променлива, която влияе върху чувствителността към чернодробно увреждане и дори кетогенезата (Garbow et al., 5; Jornayvaz et al., 2011; Kennedy et al., 2010; Pissios et al., 2007; Schugar и др., 2013). Ефектите от дългосрочната консумация на кетогенни диети при мишки остават непълно дефинирани, но последните проучвания при мишки разкриват нормална преживяемост и липса на маркери за увреждане на черния дроб при мишки на кетогенни диети през целия им живот, въпреки че метаболизмът на аминокиселините, разходът на енергия и инсулиновата сигнализация бяха значително препрограмирани (Douris et al., 2013).
Механизмите, повишаващи кетозата чрез механизми, алтернативни на кетогенните диети, включват използването на поглъщани прекурсори на кетонни тела. Приложението на екзогенни кетонни тела може да създаде уникално физиологично състояние, което не се среща в нормалната физиология, тъй като концентрациите на циркулираща глюкоза и инсулин са относително нормални, докато клетките могат да спестят усвояването и използването на глюкоза. Самите кетонни тела имат кратък полуживот и поглъщането или вливането на натриева ?OHB сол за постигане на терапевтична кетоза провокира неблагоприятно натоварване с натрий. R/S-1,3-бутандиолът е нетоксичен диалкохол, който лесно се окислява в черния дроб, за да се получи d/l-?OHB (Desrochers et al., 1992). В различни експериментални контексти тази доза е прилагана ежедневно на мишки или плъхове в продължение на цели седем седмици, което води до циркулиращи концентрации на ?OHB до 5 mM в рамките на 2 часа след приложението, което е стабилно за поне допълнителни 3 часа (D' Агостино и др., 2013). Частично потискане на приема на храна е наблюдавано при гризачи, получаващи R/S-1,3-бутандиол (Carpenter and Grossman, 1983). В допълнение, три химически различни кетонни естера (KEs), (i) моноестер на R-1,3-бутандиол и d-pOHB (R-3-хидроксибутил R-pOHB); (ii) глицерил-трис-аОНВ; и (iii) R,S-1,3-бутандиол ацетоацетат диестер също са подробно изследвани (Brunengraber, 1997; Clarke et al., 2012a; Clarke et al., 2012b; Desrochers et al., 1995a; Desrochers et al. ., 1995b; Kashiwaya et al., 2010). Присъщо предимство на първото е, че 2 мола физиологичен d-?OHB се произвеждат на мол KE, след хидролиза на естераза в червата или черния дроб. Безопасността, фармакокинетиката и толерантността са най-обстойно проучени при хора, поглъщащи R-3-хидроксибутил R-?OHB, в дози до 714 mg/kg, което води до циркулиращи концентрации на d-?OHB до 6 mM (Clarke et al., 2012a; Cox et al., 2016; Kemper et al., 2015; Shivva et al., 2016). При гризачи този KE намалява приема на калории и общия холестерол в плазмата, стимулира кафявата мастна тъкан и подобрява инсулиновата резистентност (Kashiwaya et al., 2010; Kemper et al., 2015; Veech, 2013). Последните открития показват, че по време на тренировка при тренирани спортисти, поглъщането на R-3-хидроксибутил R-?OHB намалява гликолизата на скелетните мускули и плазмените концентрации на лактат, повишава интрамускулното окисление на триацилглицерол и запазва съдържанието на мускулен гликоген, дори при едновременно поглъщане на инсулинова секреция (стимулирана секреция на инсулин Кокс и др., 2016). Необходимо е по-нататъшно развитие на тези интригуващи резултати, тъй като подобрението в изпълнението на упражненията за издръжливост се дължи предимно на силен отговор на KE при 2/8 субекта. Независимо от това, тези резултати подкрепят класическите проучвания, които показват предпочитание към окисление на кетон пред други субстрати (GARLAND et al., 1962; Hasselbaink et al., 2003; Stanley et al., 2003; Valente-Silva et al., 2015), включително по време на тренировка и че тренираните спортисти могат да бъдат по-подготвени да използват кетони (Johnson et al., 1969a; Johnson and Walton, 1972; Winder et al., 1974; Winder et al., 1975). И накрая, механизмите, които биха могли да подкрепят подобрената производителност на упражненията след равен прием на калории (диференциално разпределени между макронутриентите) и равни нива на консумация на кислород остават да бъдат определени.
Бъдеща перспектива
Веднъж до голяма степен стигматизиран като преливащ път, способен да натрупва токсични емисии от изгарянето на мазнини в състояния с ограничени въглехидрати (парадигмата на „кетотоксичната“), последните наблюдения подкрепят схващането, че метаболизмът на кетоновите тела изпълнява благотворна роля дори в състояния, натоварени с въглехидрати, отваряйки „кетохорметик“. хипотеза. Докато лесните хранителни и фармакологични подходи за манипулиране на метаболизма на кетоните го правят привлекателна терапевтична цел, агресивно поставени, но разумни експерименти остават както в основните, така и в транслационните изследователски лаборатории. Появиха се неудовлетворени нужди в областите на определяне на ролята на усвояването на кетонния метаболизъм при сърдечна недостатъчност, затлъстяване, NAFLD/NASH, диабет тип 2 и рак. Обхватът и въздействието на „неканоничните“ сигнални роли на кетонните тела, включително регулирането на PTMs, които вероятно се връщат назад и напред в метаболитни и сигнални пътища, изискват по-задълбочено изследване. И накрая, екстрахепаталната кетогенеза може да отвори интригуващи паракринни и автокринни сигнални механизми и възможности за повлияване на съвместния метаболизъм в нервната система и туморите за постигане на терапевтични цели.
В заключение, кетонните тела се създават от черния дроб, за да се използват като източник на енергия, когато в човешкото тяло няма достатъчно глюкоза. Кетогенезата възниква, когато има ниски нива на глюкоза в кръвта, особено след изчерпване на други клетъчни запаси от въглехидрати. Целта на статията по-горе беше да се обсъдят многоизмерните роли на кетонните тела в горивния метаболизъм, сигнализирането и терапията. Обхватът на нашата информация е ограничен до хиропрактика и проблеми със здравето на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Допълнителна дискусия на тема: „Остра болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати работни дни в световен мащаб. Болката в гърба се приписва на втората най-честа причина за посещения на лекар, превъзхождаща само инфекциите на горните дихателни пътища. Приблизително 80 процента от населението ще изпита болки в гърба поне веднъж през живота си. Гръбначният стълб е сложна структура, съставена от кости, стави, връзки и мускули, наред с други меки тъкани. Наранявания и/или влошени състояния, като напрхерния дискове, в крайна сметка може да доведе до симптоми на болки в гърба. Спортните наранявания или нараняванията при автомобилни катастрофи често са най-честата причина за болки в гърба, но понякога най-простите движения могат да имат болезнени резултати. За щастие, алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
- ядрен еритроиден 2-свързан фактор 2 сигнален път, най-известен като Nrf2, е защитен механизъм, който функционира като "главен регулатор" на антиоксидантния отговор на човешкото тяло. Nrf2 усеща нивата на оксидативен стрес в клетките и задейства защитните антиоксидантни механизми. Докато активирането на Nrf2 може да има много предимства, Nrf2 "свръхекспресия" може да има няколко рискове.
Изглежда, че балансираната степен на NRF2 е от съществено значение за предотвратяване на цялостното развитие на различни заболявания в допълнение към общото подобрение на тези здравни проблеми. Въпреки това NRF2 може да причини усложнения. Основната причина за "свръхекспресията" на NRF2 се дължи на генетична мутация или продължаващо хронично излагане на химически или оксидативен стрес, между другото. По-долу ще обсъдим недостатъците на свръхекспресията на Nrf2 и ще демонстрираме нейните механизми на действие в човешкото тяло.
Рак
Изследователски изследвания установиха, че мишките, които не експресират NRF2, са по-склонни да развият рак в отговор на физическа и химическа стимулация. Подобни изследователски проучвания обаче показват, че прекомерното активиране на NRF2 или дори инактивирането на KEAP1 може да доведе до обостряне на някои видове рак, особено ако тези пътища са били прекъснати. Свръхактивността на NRF2 може да възникне чрез пушене, където се смята, че продължителното активиране на NRF2 е причината за рак на белия дроб при пушачите. Свръхекспресията на Nrf2 може да доведе до саморазрушаване на раковите клетки, докато периодичното активиране на NRF2 може да попречи на раковите клетки да предизвикат индукция на токсин.
Освен това, тъй като свръхекспресията на NRF2 увеличава антиоксидантната способност на човешкото тяло да функционира извън редокс хомеостазата, това засилва клетъчното делене и генерира неестествен модел на ДНК и метилиране на хистон. Това в крайна сметка може да направи химиотерапията и лъчетерапията по-малко ефективни срещу рак. Следователно ограничаването на активирането на NRF2 с вещества като DIM, Luteolin, Zi Cao или salinomycin може да бъде идеално за пациенти с рак, въпреки че свръхактивирането на Nrf2 не трябва да се счита за единствената причина за рак. Недостигът на хранителни вещества може да повлияе на гените, включително NRF2. Това може да е един от начините за това как дефицитите допринасят за туморите.
Черен дроб
Преактивирането на Nrf2 може също да повлияе на функцията на определени органи в човешкото тяло. Прекомерната експресия на NRF2 може в крайна сметка да блокира производството на инсулиноподобен растежен фактор 1 или IGF-1 от черния дроб, което е от съществено значение за регенерацията на черния дроб.
NXNUMX Сърце
Докато острата свръхекспресия на Nrf2 може да има своите предимства, непрекъснатото свръхекспресия на NRF2 може да причини дълготрайни вредни ефекти върху сърцето, като кардиомиопатия. NRF2 експресията може да бъде увеличена чрез високи нива на холестерол или чрез активиране на HO-1. Това се смята за причината, поради която хроничните повишени нива на холестерол могат да причинят сърдечносъдови проблеми.
Витилиго
Известно е, че свръхекспресията на NRF2 инхибира способността за повторно потискане на витилиго, тъй като може да възпрепятства тирозиназата или TYR, действие, което е от съществено значение за репагментацията чрез меланиногенеза. Проучванията показват, че този процес може да бъде една от основните причини защо хората с витилиго не изглежда да активират Nrf2 толкова ефективно, колкото хората без витилиго.
Защо NRF2 може да не функционира правилно
хормезис
NRF2 трябва да бъде хормонално активиран, за да може да се възползва от предимствата му. С други думи, Nrf2 не трябва да се задейства всяка минута или всеки ден, следователно е страхотна идея да правите паузи от него, например 5 дни на 5 почивни дни или през ден. NRF2 също трябва да постигне специфичен праг, за да задейства своя хормонален отговор, където малък стресор може да не е достатъчен, за да го задейства.
DJ-1 Окисление
Протеинът deglycase DJ-1, или просто DJ-1, наречен протеин на болестта на Паркинсон или PARK7, е главен регулатор и детектор на редукционния статус в човешкото тяло. DJ-1 е от съществено значение за регулиране колко дълго NRF2 може да изпълнява своята функция и да произвежда антиоксидантен отговор. В случай, че DJ-1 се преокислява, клетките ще направят DJ-1 протеина по-малко достъпен.
Този процес индуцира активирането на NRF2 да изтече твърде бързо, тъй като DJ-1 е от първостепенно значение за поддържането на балансирани нива на NRF2 и предотвратяването им от разпадането им в клетката. В случай, че протеинът DJ-1 е несъществуващ или преоксидиран, експресията на NRF2 вероятно ще бъде минимална дори при използване на DIM или алтернативни NRF2 активатори. Изразяването на DJ-1 е наложително за възстановяване на нарушеното действие на NRF2.
Хронично заболяване
Ако имате хронично заболяване, включително CIRS, хронични инфекции/дисбиоза/SIBO или натрупване на тежки метали, като живак и/или такива от кореновите канали, те могат да възпрепятстват системите на NRF2 и фаза втора детоксикация. Вместо оксидативният стрес да превръща NRF2 в антиоксидант, NRF2 няма да се задейства и оксидативният стрес може да остане в клетката и да причини увреждане, което означава, че няма антиоксидантен отговор. Това е значителна причина, поради която много хора с CIRS имат няколко чувствителност и достигат до множество фактори. Някои хора вярват, че може да имат реакция на херкс, но тази реакция може само да уврежда клетките по-далеч.
Лечението на хронично заболяване обаче ще позволи на черния дроб да изхвърля токсините в жлъчката, като постепенно развива хормоналния отговор на активиране на NRF2. Ако жлъчката остане токсична и не се екскретира от човешкото тяло, тя ще реактивира оксидативния стрес на NRF2 и ще ви накара да се почувствате по-зле, след като се резорбира от стомашно-чревния или стомашно-чревния тракт. Например, охратоксин А може да блокира NRF2. Освен че лекуват проблема, инхибиторите на хистон деацетилаза могат да блокират окислителната реакция от редица фактори, които предизвикват активиране на NRF2, но могат също така да попречат на NRF2 да се задейства – нормално, което в крайна сметка може да не изпълни целта си.
Регулиране на рибеното масло
Холинергичните вещества са вещества, които усилват ацетилхолин или ACh и холин в мозъка чрез увеличаване на ACh, особено когато инхибират разграждането на ACh. Пациентите с CIRS често имат проблеми с нарушаването на нивата на ацетилхолина в човешкото тяло, особено в мозъка. Рибеното масло задейства NRF2, активирайки защитния му антиоксидантен механизъм в клетките.
Хората с хронични заболявания може да имат проблеми с когнитивния стрес и ацетилхолиновата ексайтотоксичност, поради натрупване на органофосфат, което може да причини рибено масло да създаде възпаление в човешкото тяло. Дефицитът на холин допълнително предизвиква активиране на NRF2. Включването на холин във вашата диета (полифеноли, яйца и др.) може да помогне за засилване на ефектите от холинергичната дисрегулация.
Какво намалява NRF2?
Намаляването на свръхекспресията с NRF2 е най-доброто за хора, които имат рак, въпреки че може да е от полза за редица други здравни проблеми.
Диета, добавки и общи лекарства:
Апигенин (по-високи дози)
Brucea javanica
кестени
EGCG (високи дози увеличават NRF2)
Сминдух (тригонелин)
Hiba (Hinokitiol /? -Туяплицин)
Висока солева диета
Лутеолин (целина, зелен пипер, магданоз, перлил и чай от лайка - по-високи дози могат да увеличат NRF2 - 40 mg / kg лутеолин три пъти седмично)
Метформин (хроничен прием)
N-Ацетил-L-цистеин (NAC, чрез блокиране на окислителния отговор, esp при високи дози)
Orange Peel (имат полиметоксилирани флавоноиди)
Кверцетин (по-високи дози могат да повишат NRF2 - 50 mg / kg / ден кверцетин)
Салиномицин (лекарство)
Ретинол (изцяло транс-ретинова киселина)
Витамин С, когато се комбинира с кверцетин
Зи Као (Purple Gromwel има Shikonin / Alkannin)
Пътища и други:
Bach1
ЗАЛАГАНЕ
Биофилми
Brusatol
капмтотецин
DNMT
DPP-23
EZH2
Сигнализиране на глюкокортикоидния рецептор (дексаметазон и бетаметазон)
GSK-3? (регулаторна обратна връзка)
HDAC активиране?
Halofuginone
Хомоцистеин (ALCAR може да обърне този хомоцистеин, предизвиква ниски нива на NRF2)
IL-24
Keap1
MDA-7
NF? B
Охратоксин А (видове аспергилус и пенциклий)
Промиелоцитна левкемия протеин
p38
p53
p97
Ретиноева киселина рецептор алфа
селенит
SYVN1 (Hrd1)
STAT3 инхибиране (като например Cryptotanshinone)
Тестостерон (и тестостерон пропионат, въпреки че TP интраназално може да увеличи NRF2)
Трактор (Етионамид)
Trx1 (чрез редукция на Cys151 в Keap1 или на Cys506 в NLS областта на Nrf2)
тролокс
Vorinostat
Цинков дефицит (влошава в мозъка)
Nrf2 Механизъм на действие
Оксидативният стрес се задейства чрез CUL3, където NRF2 от KEAP1, отрицателен инхибитор, след това навлиза в ядрото на тези клетки, стимулира транскрипцията на AREs, превръща сулфидите в дисулфиди и ги превръща в по-антиоксидантни гени, водещи до повишаване на регулирането на антиоксиданти като GSH, GPX, GST, SOD и др. Останалите от тях могат да се видят в списъка по-долу:
Увеличава AKR
Увеличава се
Увеличава ATF4
Увеличава Bcl-xL
Увеличава Bcl-2
Увеличава BDNF
Увеличава BRCA1
Увеличава c-Jun
Увеличава CAT
Увеличава cGMP
Увеличава CKIP-1
Увеличава CYP450
Увеличава Cul3
Увеличава GCL
Увеличава GCLC
Увеличава GCLM
Увеличава GCS
Увеличава GPx
Увеличава GR
Увеличава GSH
Увеличава GST
Увеличава HIF1
Увеличава HO-1
Увеличава HQO1
Увеличава HSP70
Увеличава IL-4
Увеличава IL-5
Увеличава IL-10
Увеличава IL-13
Увеличава K6
Увеличава K16
Увеличава K17
Увеличава mEH
Увеличава Mrp2-5
Увеличава NADPH
Увеличава Notch 1
Увеличава NQO1
Увеличава PPAR-алфа
Увеличава Prx
Увеличава p62
Увеличава Sesn2
Увеличава Slco1b2
Увеличава sMafs
Увеличава SOD
Увеличава Trx
Увеличава Txn (d)
Увеличава UGT1 (A1 / 6)
Увеличава VEGF
Намалява ADAMTS (4 / 5)
Намалява алфа-SMA
Намалява ALT
Намалява AP1
Намалява AST
Намалява Bach1
Намалява COX-2
Намалява DNMT
Намалява FASN
Намалява FGF
Намалява HDAC
Намалява IFN-?
Намалява IgE
Намалява IGF-1
Намалява IL-1b
Намалява IL-2
Намалява IL-6
Намалява IL-8
Намалява IL-25
Намалява IL-33
Намалява iNOS
Намалява LT
Намалява Keap1
Намалява MCP-1
Намалява MIP-2
Намалява MMP-1
Намалява MMP-2
Намалява MMP-3
Намалява MMP-9
Намалява MMP-13
Намалява NfkB
Намалява НЕ
Намалява SIRT1
Намалява TGF-b1
Намалява TNF-алфа
Намалява Tyr
Намалява VCAM-1
Кодиран от ген NFE2L2, NRF2 или ядрен еритроиден 2-свързан фактор 2, е транскрипционен фактор в основния левцин цип или bZIP, суперфамилия, която използва структура Cap'n'Collar или CNC.
Той подпомага азотните ензими, биотрансформационните ензими и ксенобиотичните ефлуксни транспортери.
Той е основен регулатор при индуцирането на антиоксидантни и детоксикационни ензимни гени от фаза II, които предпазват клетките от увреждане, причинено от оксидативен стрес и електрофилни атаки.
При хомеостатични условия Nrf2 се изолира в цитозола чрез телесно прикрепване на N-терминалния домен на Nrf2 или Kelch-подобния ECH-свързан протеин или Keap1, също така наречен INrf2 или инхибитор на Nrf2, инхибиращ Nrf2 активиране.
Той може също да бъде контролиран от селенопротеин тиореоксин редуктаза от бозайник 1 или TrxR1, който функционира като отрицателен регулатор.
При уязвимостта към електрофилните стресиращи средства, Nrf2 се разпада от Keap1, пренасяйки се в ядрото, където след това се хетеродимеризира с редица транскрипционни регулаторни протеини.
Честите взаимодействия се състоят от тези на транскрипционните органи Jun и Fos, които могат да бъдат членове на фамилията на транскрипционните фактори на протеиновия активатор.
След димеризацията, тези комплекси се свързват с анти-оксидант / електрофилен отзивчив компонент ARE / EpRE и активират транскрипцията, както е вярно при Jun-Nrf2 комплекса, или потискат транскрипцията, подобно на Fos-Nrf2 комплекс.
Позиционирането на ARE, което се задейства или инхибира, ще определи кои гени са транскрипционно контролирани от тези променливи.
Когато ARE се задейства:
Активирането на синтеза на антиоксиданти е способно да детоксикира ROS като каталаза, супероксид-дисмутаза или SOD, GSH-пероксидази, GSH-редуктаза, GSH-трансфераза, NADPH-хинон оксидоредуктаза или NQO1, цитохром Р450, моноокситиогеназа, моноокситиогеназа, система редуктаза и HSP70.
Активирането на тази GSH синтаза позволява забележим растеж на вътреклетъчната степен на GSH, което е доста защитно.
Увеличаването на този синтез и степените на ензими от фаза II като UDP-глюкуронозилтрансфераза, N-ацетилтрансферази и сулфотрансферази.
Повишената регулация на HO-1, която е наистина защитен рецептор с потенциален растеж на СО, който заедно с NO позволява вазодилатация на исхемични клетки.
Намаляване на претоварването с желязо чрез повишен феритин и билирубин като липофилен антиоксидант. Както протеините фаза II, така и антиоксидантите са в състояние да определят хроничния оксидативен стрес и да възстановят нормалната редукционна система.
GSK3? под управлението на AKT и PI3K, фосфорилира Fyn, което води до ядрена локализация на Fyn, която Fyn фосфорилира Nrf2Y568, което води до ядрен износ и разграждане на Nrf2.
NRF2 също така смекчава реакцията TH1 / TH17 и обогатява отговора TH2.
HDAC инхибиторите задействат сигналната пътека Nrf2 и се регулират нагоре, че Nrf2 надолу по веригата каталитична субединица HO-1, NQO1 и глутамат-цистеин лигаза или GCLC чрез ограничаване на Keap1 и насърчаване на дисоциацията на Keap1 от Nrf2, Nrf2 ядрена транслокация и Nrf2 -ARE задължително.
Nrf2 включва период на полуразпад от около 20 минути при базови условия.
Намаляване на IKK? пул чрез свързване Keap1 намалява I? B? деградация и може да бъде неуловим механизъм, чрез който е доказано, че активирането на Nrf2 инхибира активирането на NF? B.
Keap1 не винаги трябва да бъде регулиран надолу, за да работи с NRF2, като хлорофилин, боровинка, елагична киселина, астаксантин и чаени полифеноли може да увеличи NRF2 и KEAP1 при 400 процента.
Nrf2 регулира отрицателно чрез термина стеароил СоА десатураза или SCD и цитратна лиаза или CL.
генетика
KEAP1
rs1048290
C алел - показа значителен риск за и защитен ефект срещу резистентна към лекарства епилепсия (DRE)
rs11085735 (аз съм AC)
свързана с степента на спадане на белодробната функция в LHS
сайтаОУСведения
rs242561
Т алел - защитен алел за Паркинсонови заболявания - има по-силно свързване с NRF2 / sMAF и е свързано с по-високите нива на MAPT mRNA в различни региони на 3 в мозъка, включително мозъчна кора (CRBL), темпорален кортекс (TCTX)
NFE2L2 (NRF2)
rs10183914 (аз съм CT)
Т алел - повишени нива на Nrf2 протеин и забавена възраст на началото на болестта на Паркинсон с четири години
rs16865105 (аз съм AC)
C алел - имаше по-висок риск от болестта на Паркинсон
rs1806649 (аз съм CT)
C алел - е идентифициран и може да бъде от значение за етиологията на рака на гърдата.
свързани с повишен риск от приемане в болница в периоди с високи нива на PM10
rs1962142 (Аз съм GG)
Т алел - е свързано с ниско ниво на цитоплазмена експресия на NRF2 (P = 0.036) и отрицателна сулфиредоксинова експресия (P = 0.042)
Алел - защитено от спадане на кръвообращението на предмишницата (FEV) (принудителен издишващ обем за една секунда) във връзка със състоянието на пушенето на цигари (p = 0.004)
rs2001350 (Аз съм TT)
Т алел - защитени от спадане на FEV (принудителен издишващ обем за една секунда) във връзка със състоянието на пушенето на цигари (p = 0.004)
rs2364722 (Аз съм АА)
Алел - защитени от спад на FEV (принудителен издишвателен обем за една секунда) във връзка със състоянието на пушенето на цигари (p = 0.004)
rs2364723
C алел - свързано със значително намаление на FEV при японски пушачи с рак на белия дроб
rs2706110
G алел - показва значителен риск за и защитен ефект срещу резистентна към лекарства епилепсия (DRE)
AA алелите - показаха значително намалено изразяване на KEAP1
AA алелите - се свързва с повишен риск от рак на гърдата (P = 0.011)
rs2886161 (Аз съм TT)
Т алел - свързан с болестта на Паркинсон
rs2886162
Алел – е свързан с ниска експресия на NRF2 (P = 0.011; OR, 1.988; CI, 1.162 3.400) и AA генотипът е свързан с по-лоша преживяемост (P = 0.032; HR, 1.687; CI, 1.047 2.748)
rs35652124 (Аз съм TT)
Алел - свързан с по-висок, свързан с възрастта при настъпване на болестта на Паркинсон срещу G алел
C алел - увеличава протеина NRF2
Т алел - има по-малък протеин NRF2 и по-голям риск от сърдечни заболявания и кръвно налягане
rs6706649 (аз съм CC)
C алел - имат по-нисък протеин NRF2 и повишават риска от болестта на Паркинсон
rs6721961 (Аз съм GG)
Т алел - имаше по-нисък протеин NRF2
ТТ алели - връзка между тютюнопушенето при тежки пушачи и намаляване на качеството на спермата
TT алел – се свързва с повишен риск от рак на гърдата [P = 0.008; OR, 4.656; доверителен интервал (CI), 1.350 × 16.063] и алелът T е свързан с ниска степен на експресия на NRF2 протеин (P = 0.0003; OR, 2.420; CI, 1.491 × 3.926) и отрицателна експресия на SRXN1 (P = 0.047; OR); 1.867; CI = 1.002 3.478)
Т алела - алелът също е номинално свързан със смъртността, свързана с ALI 28-ден, след синдром на системната възпалителна реакция
Т алел - защитени от спадане на FEV (принудителен издишващ обем за една секунда) във връзка със състоянието на пушенето на цигари (p = 0.004)
G алел - свързан с повишен риск от ALI след тежка травма в Европа и Африка-Америка (коефициент на вероятност, или 6.44; 95% доверителен интервал
AA алели - свързани с инфекция-индуцирана астма
AA алелите - проявяват значително намалена експресия на ген NRF2 и следователно повишен риск от рак на белия дроб, особено тези, които някога са пушили
АА-алелите - има значително по-висок риск от развитие на T2DM (OR 1.77; 95% CI 1.26, 2.49; p = 0.011) спрямо тези с CC генотип
AA алели - силна асоциация между ремонта на рани и късната токсичност на радиацията (свързана със значително по-висок риск за развитие на късни ефекти в афро-американците с тенденция към кавказки)
свързани с перорална естрогенна терапия и риск от венозен тромбоемболизъм при жени в менопауза
rs6726395 (Аз съм AG)
Алел - защитено от спадане на FEV1 (принудително издишване за една секунда) във връзка със състоянието на пушенето на цигари (p = 0.004)
Алел - свързан със значително намален FEV1 при японски пушачи с рак на белия дроб
GG алели - имат по-високи нива на NRF2 и намален риск от макулна дегенерация
GG алели - имат по-висока преживяемост с холангиокарцином
rs7557529 (аз съм CT)
C алел - свързан с болестта на Паркинсон
Оксидативният стрес и други стресови фактори могат да причинят увреждане на клетките, което в крайна сметка може да доведе до различни здравни проблеми. Изследванията показват, че активирането на Nrf2 може да насърчи защитния антиоксидантен механизъм на човешкото тяло, обаче изследователите обсъдиха, че свръхекспресията на Nrf2 може да има огромни рискове за цялостното здраве и уелнес. Различни видове рак могат да възникнат и при свръх активиране на Nrf2.
Д-р Алекс Хименес DC, CCST Insight
Сулфорафан и неговите ефекти върху рака, смъртността, стареенето, мозъка и поведението, сърдечните заболявания и др.
Изотиоцианатите са едни от най-важните растителни съединения, които можете да получите във вашата диета. В това видео правя най-изчерпателния случай за тях, който някога е бил направен. Кратко внимание? Прескочи до любимата си тема, като кликнете върху някоя от точките по-долу. Пълна времева линия по-долу.
Ключови секции:
00: 01: 14 - Рак и смъртност
00: 19: 04 - Стареене
00: 26: 30 - Мозък и поведение
00: 38: 06 - Краен рекап
00: 40: 27 - Доза
Пълна времева линия:
00: 00: 34 - Въвеждане на сулфорафан, основен фокус на видеото.
00: 01: 14 - Консумация на кръстоцветни зеленчуци и намаляване на смъртността от всякаква причина.
00: 02: 12 - риск от рак на простатата.
00: 02: 23 - риск от рак на пикочния мехур.
00: 02: 34 - Рак на белия дроб при риск от пушене.
00: 02: 48 - риск от рак на гърдата.
00: 03: 13 - Хипотетично: какво ще стане, ако вече имате рак? (Интервенция)
00: 03: 35 - Вероятен механизъм, който управлява асоциативните данни за рака и смъртността.
00: 04: 38 - Сулфорафан и рак.
00: 05: 32 - доказателства при животни, показващи силен ефект на екстракт от кълнове от броколи върху развитието на тумор на пикочния мехур при плъхове.
00: 06: 06 - Ефект на директно добавяне на сулфорафан при пациенти с рак на простатата.
00: 07: 09 - Биоакумулиране на изотиоцианатни метаболити в действителната гръдна тъкан.
00: 08: 32 - Инхибиране на стволови клетки от рак на гърдата.
00: 08: 53 - История: Brassicas са установени със здравни качества дори в древния Рим.
00: 09: 16 - Способността на Sulforaphane да подобри екскрецията на канцероген (бензен, акролеин).
00: 09: 51 - NRF2 като генетичен превключвател чрез елементи на антиоксидантен отговор.
00: 10: 10 - Как активирането на NRF2 подобрява екскрецията на канцероген чрез глутатион-S-конюгати.
00: 10: 34 - Брюкселското зеле увеличава глутатион-8-трансферазата и намалява увреждането на ДНК.
00: 11: 20 - Напитката от кълнове от броколи повишава екскрецията на бензен с 61%.
00: 15: 45 - Кръстосана консумация на зеленчуци и смъртност от сърдечни заболявания.
00: 16: 55 - Броколи зеленчуков прах подобрява кръвните липиди и цялостния риск от сърдечни заболявания при диабетици тип 2.
00: 19: 04 - начало на стареене секция.
00: 19: 21 - Диета, обогатена със сулфарафан, увеличава живота на бръмбарите от 15 до 30% (при определени условия).
00: 20: 34 - Значение на ниското възпаление за дълголетие.
00: 22: 05 - Кръстоцветните зеленчуци и кълнове от кълнове от броколи изглежда намаляват голямо разнообразие от възпалителни маркери при хората.
00: 23: 40 - Средно видеоклип: рак, стареене
00: 24: 14 - Изследванията на мишки показват, че сулфарафан може да подобри адаптивната имунна функция в напреднала възраст.
00: 25: 18 - Сулфарафан подобри растежа на косата при миши модел на оплешивяване. Изображение на 00: 26: 10.
00: 26: 30 - начало на секцията за мозъка и поведението.
00: 27: 18 - Ефект на екстракт от кълнове от броколи върху аутизма.
00: 27: 48 - Ефект на глюкорафанин върху шизофрения.
00: 28: 17 - Начало на депресивната дискусия (правдоподобен механизъм и проучвания).
00: 31: 21 - Изследване на мишки, използващи различни модели 10 на стресова индуцирана депресия, показва сулфорафан с подобен ефект като флуоксетин (prozac).
00: 32: 00 - Изследването показва, че директното поглъщане на глюкорафанин в мишки е подобно ефективно при предотвратяване на депресията от модела на социално потискане на стреса.
00: 33: 01 - начало на секцията за невродегенерация.
00: 33: 30 - Сулфорафан и болестта на Алцхаймер.
00: 33: 44 - Сулфорафан и болестта на Паркинсон.
00: 33: 51 - Сулфорафан и болестта на Хънтингтън.
00: 34: 13 - Сулфорафан увеличава протеините на топлинния шок.
00: 34: 43 - Начало на травматичното мозъчно увреждане.
00: 35: 01 - Сулфарафан се инжектира веднага след като TBI подобрява паметта (изследване на мишката).
00: 35: 55 - Сулфорафан и невронна пластичност.
00: 36: 32 - Сулфорафан подобрява ученето при модел на диабет тип II при мишки.
00: 37: 19 - Сулфорафан и душевна мускулна дистрофия.
00: 37: 44 - Инхибиране на миостатина в мускулните сателитни клетки (in vitro).
00: 38: 06 - Късно видеоклип: смъртност и рак, увреждане на ДНК, оксидативен стрес и възпаление, екскреция на бензол, сърдечно-съдови заболявания, диабет тип II, ефекти върху мозъка (депресия, аутизъм, шизофрения, невродегенерация).
00: 40: 27 - Мисли за определяне на доза от кълнове от броколи или сулфорафан.
00: 41: 01 - Анекдоти за поникване у дома.
00: 43: 14 - На температурата на готвене и активността на сулфорафан.
00: 43: 45 - Конверсия на сулфорафан от глюкорафанин в чревната бактерия.
00: 44: 24 - Добавките работят по-добре, когато се комбинират с активна мирозиназа от зеленчуци.
00: 44: 56 - техники за готвене и кръстоцветни зеленчуци.
00: 46: 06 - Изотиоцианати като глюкогени.
Според изследвания, Nrf2 е основен транскрипционен фактор, който активира защитните антиоксидантни механизми на клетките за детоксикация на човешкото тяло. Свръхекспресията на Nrf2 обаче може да причини здравословни проблеми. Обхватът на нашата информация е ограничен до хиропрактика и проблеми със здравето на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителна дискусия на тема: „Остра болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати работни дни в световен мащаб. Болката в гърба се приписва на втората най-честа причина за посещения на лекар, превъзхождаща само инфекциите на горните дихателни пътища. Приблизително 80 процента от населението ще изпита болки в гърба поне веднъж през живота си. Гръбначният стълб е сложна структура, съставена от кости, стави, връзки и мускули, наред с други меки тъкани. Наранявания и/или влошени състояния, като напрхерния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Много настоящи изследвания върху рака позволиха на здравните специалисти да разберат начина, по който тялото се детоксикира. Чрез анализиране на регулирани гени в туморни клетки, изследователите откриха ядрен еритроиден 2-свързан фактор 2 сигнален път, най-известен като Nrf2. NRF2 е важен транскрипционен фактор, който активира човешкия организъм защитни антиоксидантни механизми с цел регулиране на окисляването както от външни, така и от вътрешни фактори, за да се предотвратят повишени нива на оксидативен стрес.
Принципи на Nrf2
NRF2 е от съществено значение за поддържането на цялостното здраве и благополучие, защото служи на основната цел да регулира как управляваме всичко, на което сме изложени ежедневно, и да не се разболеем. Активирането на NRF2 играе роля в системата за детоксикация от фаза II. Детоксикацията от фаза II отнема липофилни или мастноразтворими свободни радикали и ги превръща в хидрофилни или водоразтворими вещества за екскреция, като в резултат на това инактивира изключително реактивни метаболити и химикали от фаза I.
Активирането на NRF2 намалява цялостното окисляване и възпаление на човешкото тяло чрез хорметичен ефект. За да се задейства NRF2, трябва да възникне възпалителна реакция, дължаща се на окисление, за да могат клетките да произведат адаптивен отговор и да създадат антиоксиданти, като глутатион. За да разруши принципа на Nrf2, по същество оксидативният стрес активира NRF2, който след това активира антиоксидантен отговор в човешкото тяло. NRF2 функционира за балансиране на редокс сигнализирането или равновесието на нивата на оксидант и антиоксидант в клетката.
Страхотна илюстрация за това как функционира този процес може да бъде демонстрирана с упражнения. Чрез всяка тренировка мускулът се адаптира, така че да може да побере друга тренировка. Ако NRF2 стане недостатъчно или свръхекспресиран поради хронични инфекции или повишена експозиция на токсини, което може да се наблюдава при пациенти със синдром на хроничен възпалителен отговор или CIRS, здравословните проблеми могат да се влошат след активиране на NRF2. Преди всичко, ако DJ-1 стане прекомерно окислен, активирането на NRF2 ще приключи твърде бързо.
Ефекти от активирането на NRF2
Активирането на NRF2 е силно изразено в белите дробове, черния дроб и бъбреците. Фактор 2, свързан с ядрен еритроид 2, или NRF2, най-често функционира, като противодейства на повишените нива на окисление в човешкото тяло, което може да доведе до оксидативен стрес. Активирането на Nrf2 може да помогне за лечение на различни здравословни проблеми, но прекомерното активиране на Nrf2 може да влоши различни проблеми, които са показани по-долу.
Периодичното активиране на Nrf2 може да помогне:
Стареене (т.е. дълголетие)
Автоимунитет и цялостно възпаление (т.е. артрит, аутизъм)
Рак и химиопротекция (т.е. излагане на ЕМП)
Депресия и тревожност (т.е. PTSD)
Излагане на наркотици (алкохол, НСПВС)
Упражнение и представяне за издръжливост
Болест на червата (напр. SIBO, дисбиоза, улцерозен колит)
Сърдечна трансплантация (докато отвореният NRF2 може да е лош, NRF2 може да помогне с ремонта)
Хепатит С
Нефрит (тежки случаи)
Витилиго
Освен това, NRF2 може да помогне за действието на специфични хранителни добавки, лекарства и лекарства. Много естествени добавки също могат да помогнат за задействането на NRF2. Чрез настоящи изследователски проучвания изследователите са показали, че голям брой съединения, за които някога се е смятало, че са антиоксиданти, са наистина прооксиданти. Това е така, защото почти всички се нуждаят от NRF2, за да функционират, дори от добавки като куркумин и рибено масло. Доказано е, че какаото например генерира антиоксидантни ефекти при мишки, които притежават гена NRF2.
Начини за активиране на NRF2
В случай на невродегенеративни заболявания като болестта на Алцхаймер, болестта на Паркинсон, инсулт или дори автоимунни заболявания, вероятно е най-добре да се регулира Nrf2, но по хормонален начин. Смесването на NRF2 активатори може също да има адитивен или синергичен ефект, тъй като понякога може да бъде зависим от дозата. Най-добрите начини за увеличаване на експресията на Nrf2 са изброени по-долу:
HIST (упражнение) + CoQ10 + Sun (те синергизират много добре)
Кълнове от броколи + LLLT върху главата и червата ми
Бутират + Супер кафе + Сутрешно слънце
Акупунктура (това е алтернативен метод, може да се използва и лазерна акупунктура)
Постене
Канабидиол (CBD)
Лъвска грива + мелатонин
Алфа-липоева киселина + DIM
Пелин
Активиране на PPAR-гама
Следният изчерпателен списък, съдържащ над 350 други начина за активиране на Nrf2 чрез диета, начин на живот и устройства, пробиотици, добавки, билки и масла, хормони и невротрансмитери, лекарства/лекарства и химикали, пътища/транскрипционни фактори, както и други начини, е само кратко ръководство за това какво може да предизвика Nrf2. За краткост в тази статия сме пропуснали над 500 други храни, хранителни добавки и съединения, които могат да помогнат за активиране на Nrf2. Следните са изброени по-долу:
Диета:
Acai Бери
Алкохол (Червеното вино е по-добро, особено ако в него има тапа, тъй като протокатехичният алдехид от тапи също може да активира NRF2. Като цяло алкохолът не се препоръчва, въпреки че остър прием увеличава NRF2. Хроничният прием може да намали NRF2.
водорасли (водорасли)
ябълки
Черен чай
Бразилски триъгълни орехи
Кълнчета от броколи (и други изотиоцианати, сулфорафан, както и кръстоцветни зеленчуци като бок чой, които имат D3T)
Боровинки (0.6-10 г/ден)
моркови (фалкаринон)
кайенски пипер (капсаицин)
целина (бутилфталид)
чага (бетулин)
Чай от лайка
Чиа
Китайски картофи
арония (арония)
Шоколад (тъмен или какао)
Канела
Кафе (като хлорогенна киселина, Cafestol и Kahweol)
Cordyceps
Риба (и миди)
Лененото
Чесън
гхи (евентуално)
Джинджифил (и кардамонин)
Годжи Бери
Грейпфрут (нарингенин – 50 mg/kg/ден нарингенин)
грозде
Зелен чай
Гуава
Сърце от палма
Хиджики/Уакаме
Медена пита
киви
Бобовите растения
Лъвската мания
Махува
манго (мангиферин)
Mangosteen
Мляко (козе, краве – чрез регулиране на микробиома)
Mulberries
Зехтин (кюспе – хидрокситирозол и олеанолова киселина)
Омега 6 мастни киселини (липоксин А4)
Осандж портокали (Морин)
Гъби от стриди
папая
Фъстъци
Гълъбовият грах
Нар (пуникалагин, елагова киселина)
прополис (пиноцембрин)
Лилави сладки картофи
рамбутан (геранин)
Лук
Рейши
Rhodiola Rosea (салидрозид)
Оризови трици (циклоартенил ферулат)
Оризово грозде
Rooibos Чай
Розмарин
Градински чай
шафранка
Сусамово масло
Соя (и изофлавони, Daidzein, Genistein)
скуош
ягоди
Тартарска елда
Мащерка
домати
Боб Тонка
куркума
Wasabi
диня
Начин на живот и устройства:
Акупунктура и електроакупунктура (чрез колагенова каскада на ECM)
Упражнение (Остри упражнения като HIST или HIIT изглежда са по-полезни за предизвикване на NRF2, докато по-продължителното упражнение не предизвиква NRF2, но повишава нивата на глутатион)
Диета с високо съдържание на мазнини (диета)
Силна топлина (сауна)
Вдишване на водород и водородна вода
Хипербарна кислородна терапия
Инфрачервена терапия (като Joovv)
Интравенозен витамин С
Ketogenic диета
Озон
Пушенето (не се препоръчва – острото пушене повишава NRF2, хроничното пушене намалява NRF2. Ако решите да пушите, Светият босилек може да ви помогне да се предпазите от понижаване на NRF2)
Слънце (UVB и инфрачервено)
Пробиотици:
Bacillus subtilis (fmbJ)
Clostridium butyricum (MIYAIRI 588)
Lactobacillus brevis
Lactobacillus casei (SC4 и 114001)
Lactobacillus collinoides
Lactobacillus gasseri (OLL2809, L13-Ia и SBT2055)
Lactobacillus helveticus (NS8)
Lactobacillus paracasei (NTU 101)
Lactobacillus plantarum (C88, CAI6, FC225, SC4)
Lactobacillus rhamnosus (GG)
Добавки, билки и масла:
Ацетил-L-карнитин (ALCAR) и карнитин
Allicin
Алфа-липоева киселина
Амектофлавон
Andrographis paniculata
Агматин
апигенин
Аргининът
Артишок (цианропикрин)
Ашваганда или Индийски женшен
Астрагал
Bacopa
телешка пържола (изогемакетон)
Берберинът
Бета-кариофилен
Биден Пилоса
Масло от семена на черен кимион (тимохинон)
Boswellia
Бутеин
бутират
Канабидиол (CBD)
Каротениоиди (като бета-каротин [синергия с ликопен – 2 × 15 mg/d ликопен], фукоксантин, зеаксантин, астаксантин и лутеин)
Ресвератрол (Piceid и други фитоестрогени по същество, Knotweed)
Шипка
Палисандрово дърво
Рутин
Сапаново дърво
Сарсапарила
Saururus chinensis
SC-E1 (гипс, жасмин, женско биле, кудзу и цвете от балон)
Schisandra
Самолечение (прунела)
Шлемник (Байкалин и Вогонин)
Овчи киселец
Си У Танг
Мурсалски
Спайкнард (аралия)
спирулина
Жълт кантарион
Sulforaphane
Съдърландия
Тао Хонг Си Ву
Подобен на бик
Лоза Бог на гръмотевиците (Triptolide)
Токофероли (като витамин Е или линалоол)
Трибулус Р
Tu Si Zi
TUDCA
Витамин А (въпреки че други ретиноиди инхибират NRF2)
Витамин С (само висока доза, ниска доза инхибира NRF2)
Витекс / Целомъдрено дърво
Бял божур (Paeoniflorin от Paeonia lactiflora)
Пелин (хиспидулин и артемизинин)
Xiao Yao Wan (Свободен и лесен скитник)
Йерба Санта (Eriodictyol)
Юан Джи (Тенуигенин)
Zi Cao (ще намали NRF2 при рак)
цинк
Зизифус хинап
Хормони и невротрансмитери:
адипонектин
Адропин
Естроген (но може да намали NRF2 в гръдната тъкан)
Мелатонинът
Прогестеронът
Хинолинова киселина (в защитен отговор за предотвратяване на ексцитотоксичност)
Серотонинът
Хормони на щитовидната жлеза като Т3 (могат да увеличат NRF2 в здрави клетки, но да го намалят при рак)
Витамин D
Наркотици/лекарства и химикали:
Парацетамолът
ацетазоламид
Амлодипин
Ауранофин
Бардоксолон метил (BARD)
бензнидазол
BHA
CDDO-имидазолид
Цефтриаксон (и бета-лактамни антибиотици)
Cialis
Дексаметазон
Диприван (пропофол)
ериодиктиол
Ексендин-4
Езетимиб
Флуорид
фумарат
HNE (окислено)
идазоксан
Неорганичен арсен и натриев арсенит
JQ1 (може да инхибира и NRF2, неизвестно)
Летаирис
мелфалан
метазоламид
Метилено синьо
нифедипин
нестероидни противовъзпалителни средства
олтипаз
ИПП (като омепразол и лансопразол)
Protandim – страхотни резултати in vivo, но слаби/несъществуващи при активиране на NRF2 при хора
Пробукол
Рапамицин
резерпин
рутений
Ситаксентан
Статини (като Lipitor и Simvastatin)
Tamoxifen
Танг Луо Нин
tBHQ
Tecfidera (диметил фумарат)
THC (не толкова силен като CBD)
Теофилин
умбелиферон
Урзодеоксихолова киселина (UDCA)
Верапамил
Виагра
4-ацетоксифенол
Пътища/транскрипционни фактори:
?7 nAChR активиране
AMPK
Билирубин
CDK20
CKIP-1
CYP2E1
EAAT
Ганкирин
Гремлин
GJA1
Н-феритин фероксидаза
HDAC инхибитори (като валпроева киселина и TSA, но могат да причинят нестабилност на NRF2)
Протеини от топлинен шок
IL-17
IL-22
Клото
let-7 (сваля mBach1 РНК)
MAPK
приемници на Майкъл (повечето)
miR-141
miR-153
miR-155 (събаря и mBach1 РНК)
miR-7 (в мозъка, помага при рак и шизофрения)
Прорез 1
Оксидативен стрес (като ROS, RNS, H2O2) и електрофили
PGC-1?
PKC-делта
PPAR-гама (синергични ефекти)
Инхибиране на сигма-1 рецептора
SIRT1 (увеличава NRF2 в мозъка и белите дробове, но може да го намали като цяло)
SIRT2
SIRT6 (в черния дроб и мозъка)
SRXN1
Инхибиране на TrxR1 (също затихване или изчерпване)
Цинков протопорфирин
4-HHE
Други:
Анкафлавин
Азбест
Авицини
Bacillus amyloliquefaciens (използва се в селското стопанство)
Въглероден окис
Daphnetin
Изчерпване на глутатион (изчерпване на 80% - 90% възможно)
Gymnaster koraiensis
Хепатит С
херпес (HSV)
Индийски ясен дърво
Индиговият корен
Изосалипурпозид
Изораментин
Монасцин
Омавелоксолон (силен, известен още като RTA-408)
PDTC
Дефицит на селен (дефицитът на селен може да увеличи NRF2)
Сибирска лиственица
Софорафлаванон G
Tadehagi triquetrum
Toona sinensis (7-DGD)
Тромпетно цвете
63171 и 63179 (силни)
Сигналният път на фактор 2, свързан с ядрен еритроид 2, най-известен с акронима Nrf2, е транскрипционен фактор, който играе основна роля за регулиране на защитните антиоксидантни механизми на човешкото тяло, особено за контролиране на оксидативния стрес. Докато повишените нива на оксидативен стрес могат да активират Nrf2, неговите ефекти са изключително засилени чрез присъствието на специфични съединения. Някои храни и добавки помагат за активиране на Nrf2 в човешкото тяло, включително изотиоцианат сулфорафан от кълнове от броколи. Д-р Алекс Хименес DC, CCST Insight
Сулфорафан и неговите ефекти върху рака, смъртността, стареенето, мозъка и поведението, сърдечните заболявания и др.
Изотиоцианатите са едни от най-важните растителни съединения, които можете да получите във вашата диета. В това видео правя най-изчерпателния случай за тях, който някога е бил направен. Кратко внимание? Прескочи до любимата си тема, като кликнете върху някоя от точките по-долу. Пълна времева линия по-долу.
Ключови секции:
00: 01: 14 - Рак и смъртност
00: 19: 04 - Стареене
00: 26: 30 - Мозък и поведение
00: 38: 06 - Краен рекап
00: 40: 27 - Доза
Пълна времева линия:
00: 00: 34 - Въвеждане на сулфорафан, основен фокус на видеото.
00: 01: 14 - Консумация на кръстоцветни зеленчуци и намаляване на смъртността от всякаква причина.
00: 02: 12 - риск от рак на простатата.
00: 02: 23 - риск от рак на пикочния мехур.
00: 02: 34 - Рак на белия дроб при риск от пушене.
00: 02: 48 - риск от рак на гърдата.
00: 03: 13 - Хипотетично: какво ще стане, ако вече имате рак? (Интервенция)
00: 03: 35 - Вероятен механизъм, който управлява асоциативните данни за рака и смъртността.
00: 04: 38 - Сулфорафан и рак.
00: 05: 32 - доказателства при животни, показващи силен ефект на екстракт от кълнове от броколи върху развитието на тумор на пикочния мехур при плъхове.
00: 06: 06 - Ефект на директно добавяне на сулфорафан при пациенти с рак на простатата.
00: 07: 09 - Биоакумулиране на изотиоцианатни метаболити в действителната гръдна тъкан.
00: 08: 32 - Инхибиране на стволови клетки от рак на гърдата.
00: 08: 53 - История: Brassicas са установени със здравни качества дори в древния Рим.
00: 09: 16 - Способността на Sulforaphane да подобри екскрецията на канцероген (бензен, акролеин).
00: 09: 51 - NRF2 като генетичен превключвател чрез елементи на антиоксидантен отговор.
00: 10: 10 - Как активирането на NRF2 подобрява екскрецията на канцероген чрез глутатион-S-конюгати.
00: 10: 34 - Брюкселското зеле увеличава глутатион-8-трансферазата и намалява увреждането на ДНК.
00: 11: 20 - Напитката от кълнове от броколи повишава екскрецията на бензен с 61%.
00: 15: 45 - Кръстосана консумация на зеленчуци и смъртност от сърдечни заболявания.
00: 16: 55 - Броколи зеленчуков прах подобрява кръвните липиди и цялостния риск от сърдечни заболявания при диабетици тип 2.
00: 19: 04 - начало на стареене секция.
00: 19: 21 - Диета, обогатена със сулфарафан, увеличава живота на бръмбарите от 15 до 30% (при определени условия).
00: 20: 34 - Значение на ниското възпаление за дълголетие.
00: 22: 05 - Кръстоцветните зеленчуци и кълнове от кълнове от броколи изглежда намаляват голямо разнообразие от възпалителни маркери при хората.
00: 23: 40 - Средно видеоклип: рак, стареене
00: 24: 14 - Изследванията на мишки показват, че сулфарафан може да подобри адаптивната имунна функция в напреднала възраст.
00: 25: 18 - Сулфарафан подобри растежа на косата при миши модел на оплешивяване. Изображение на 00: 26: 10.
00: 26: 30 - начало на секцията за мозъка и поведението.
00: 27: 18 - Ефект на екстракт от кълнове от броколи върху аутизма.
00: 27: 48 - Ефект на глюкорафанин върху шизофрения.
00: 28: 17 - Начало на депресивната дискусия (правдоподобен механизъм и проучвания).
00: 31: 21 - Изследване на мишки, използващи различни модели 10 на стресова индуцирана депресия, показва сулфорафан с подобен ефект като флуоксетин (prozac).
00: 32: 00 - Изследването показва, че директното поглъщане на глюкорафанин в мишки е подобно ефективно при предотвратяване на депресията от модела на социално потискане на стреса.
00: 33: 01 - начало на секцията за невродегенерация.
00: 33: 30 - Сулфорафан и болестта на Алцхаймер.
00: 33: 44 - Сулфорафан и болестта на Паркинсон.
00: 33: 51 - Сулфорафан и болестта на Хънтингтън.
00: 34: 13 - Сулфорафан увеличава протеините на топлинния шок.
00: 34: 43 - Начало на травматичното мозъчно увреждане.
00: 35: 01 - Сулфарафан се инжектира веднага след като TBI подобрява паметта (изследване на мишката).
00: 35: 55 - Сулфорафан и невронна пластичност.
00: 36: 32 - Сулфорафан подобрява ученето при модел на диабет тип II при мишки.
00: 37: 19 - Сулфорафан и душевна мускулна дистрофия.
00: 37: 44 - Инхибиране на миостатина в мускулните сателитни клетки (in vitro).
00: 38: 06 - Късно видеоклип: смъртност и рак, увреждане на ДНК, оксидативен стрес и възпаление, екскреция на бензол, сърдечно-съдови заболявания, диабет тип II, ефекти върху мозъка (депресия, аутизъм, шизофрения, невродегенерация).
00: 40: 27 - Мисли за определяне на доза от кълнове от броколи или сулфорафан.
00: 41: 01 - Анекдоти за поникване у дома.
00: 43: 14 - На температурата на готвене и активността на сулфорафан.
00: 43: 45 - Конверсия на сулфорафан от глюкорафанин в чревната бактерия.
00: 44: 24 - Добавките работят по-добре, когато се комбинират с активна мирозиназа от зеленчуци.
00: 44: 56 - техники за готвене и кръстоцветни зеленчуци.
00: 46: 06 - Изотиоцианати като глюкогени.
Според много настоящи изследвания, сигналният път, свързан с ядрен еритроид 2 фактор 2, най-известен като Nrf2, е основен транскрипционен фактор, който активира защитните антиоксидантни механизми на клетките за детоксикация на човешкото тяло както от външни, така и от вътрешни фактори и предотвратява повишеното нива на оксидативен стрес. Обхватът на нашата информация е ограничен до хиропрактика и проблеми със здравето на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителна дискусия на тема: „Остра болка в гърба
Заболявания на опорно-двигателния апарат е една от най-разпространените причини за инвалидност и пропуснати работни дни в световен мащаб. Болката в гърба се приписва на втората най-честа причина за посещения на лекар, превъзхождаща само инфекциите на горните дихателни пътища. Приблизително 80 процента от населението ще изпита болки в гърба поне веднъж през живота си. Гръбначният стълб е сложна структура, съставена от кости, стави, връзки и мускули, наред с други меки тъкани. Наранявания и/или влошени състояния, като напрхерния дискове, в крайна сметка може да доведе до симптоми на болки в гърба. Спортните наранявания или нараняванията при автомобилни катастрофи често са най-честата причина за болки в гърба, но понякога най-простите движения могат да имат болезнени резултати. За щастие, алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Оксидативният стрес има основен принос за развитието на различни здравни проблеми, включително рак, сърдечни заболявания, диабет, ускорено стареене и невродегенерация. Богати на антиоксиданти храни, билки и добавки могат да се използват за защита на човешкото тяло от високи нива на оксидативен стрес. Последните проучвания показват, че Nrf2 генен път може да помогне за засилване на ефектите на антиоксидантите. В предимствата на Nrf2 са описани по-долу.
Предпазва тялото от токсини
NRF2 е вътрешно присъщо вещество, което може да предпази клетките от вредни вътрешни и външни съединения. NRF2 може да помогне за обогатяване на реакцията на човешкото тяло към лекарства/лекарства и токсини, подобрявайки производството на...протеини, които помагат за елиминирането на съединения от клетката, известни като протеини, свързани с множествена лекарствена резистентност, или MRPs. Например, NRF2 се задейства при вдишване на цигарен дим, за да се позволи на белите дробове да се детоксикират.
Освен това е важно белите дробове да се предпазват от алергени, вирусни заболявания, бактериални ендотоксини, хипероксия и различни замърсители на околната среда. Постоянният тригер на Nrf2 обаче може да намали нивата на вещество, известно като глутатион, в човешкото тяло. NRF2 може също да предпази черния дроб от токсичност и може да предпази черния дроб от хепатотоксичност с арсен. Освен това NRF2 предпазва черния дроб и мозъка от консумация на алкохол. Например, Nrf2 може да предпазва от токсичност на ацетаминофен.
Бори се с възпалението и оксидативния стрес
Активирането на NRF2 може да помогне в борбата срещу възпалението чрез намаляване на възпалителните цитокини, като тези, присъстващи при псориазис. NRF2 може също да намали възпалението, свързано с различни здравословни проблеми като артрит и фиброза на черния дроб, бъбреците и белите дробове. NRF2 може също да помогне за контролиране на алергиите чрез понижаване на Th1/Th17 цитокините и повишаване на TH2 цитокините. Това може да бъде от полза при заболявания като астма.
NRF2 допълнително предпазва от увреждане на клетките от синя светлина и от UVA/UVB, намиращи се на слънчева светлина. Дефицитите на Nrf2 могат да направят много по-лесно изгарянето на слънце. Една от причините за това е, че NRF2 е способен да регулира колагена в отговор на UV радиация. Усъвършенстваните крайни продукти на гликация или AGEs допринасят за развитието на много здравословни проблеми, включително диабет и невродегенеративни заболявания. NRF2 може да намали оксидативния стрес на AGEs в тялото. NRF2 може също да предпази човешкото тяло от по-високи нива на топлинен стрес.
Подобрява ефективността на митохондриите и упражненията
NRF2 е митохондриален бустер. Активирането на NRF2 допринася за повишаване на енергията на АТФ за митохондриите, в допълнение към засиленото използване на кислород или цитрат и мазнини. Без NRF2 митохондриите просто биха имали способността да функционират със захар или глюкоза, а не с мазнини. NRF2 също е от съществено значение за развитието на митохондриите чрез процес, известен като биогенеза. Активирането на NRF2 е жизненоважно, за да се „възползват“ от предимствата на упражненията.
Поради активността на Nrf2, упражненията повишават митохондриалната функция, като този резултат може да бъде усилен с CoQ10, Cordyceps и калорично ограничение. Умереното упражнение или острото упражнение предизвиква митохондриална биогенеза и повишен синтез на супероксид дисмутаза, или SOD, и хем-оксигеназа-1, или HO-1, чрез активиране на NRF2. Алфа-липоевата киселина, или ALA, и Дан Шен могат да засилят медиираната от NRF2 митохондриална биогенеза. Освен това, NRF2 може също да подобри толерантността към упражнения, където изтриването на NRF2 прави упражненията вредни.
Предпазва от хипоксия
NRF2 също помага за защита на човешкото тяло от клетъчна загуба/изчерпване на кислород, здравословен проблем, наречен хипоксия. Хората с CIRS имат намалени нива на кислород, тъй като техният NRF2 е възпрепятстван, което води до намалени нива на VEGF, HIF1 и HO-1. Обикновено при здрави индивиди с хипоксия, miR-101, който е необходим за създаването на стволови клетки, е свръхекспресиран и повишава количествата NRF2/HO-1 и VEGF/eNOS, като по този начин предотвратява увреждането на мозъка, но това не изглежда да се случи в CIRS.
Хипоксията, характеризираща се с нисък HIF1, при CIRS може също да доведе до пропусклива кръвно-мозъчна бариера поради дисбаланс на NRF2. Салидрозид, намиращ се в родиола, действа върху активирането на NRF2 и подпомага хипоксията чрез повишаване на нивата на VEGF и HIF1 в човешкото тяло. NRF2 може също така в крайна сметка да предпази от натрупване на лактат в сърцето. Активирането на NRF2 може също да спре индуцираната от хипоксия височинна болест при движение или AMS.
Забавя стареенето
Няколко съединения, които могат да бъдат фатални в огромни количества, могат да увеличат дълголетието в доста малки количества поради ксенохормеза чрез NRF2, PPAR-гама и FOXO. Много малко количество токсини повишава способността на клетката да бъде по-добре оборудвана за следващия път, когато бъде предизвикана с токсин, но това не е одобрение за консумация на отровни химикали.
Добра илюстрация на този процес е ограничението на калориите. NRF2 може да подобри продължителността на живота на клетките, като повиши техните нива на митохондриите и антиоксидантите, както и понижи способността на клетките да умрат. NRF2 намалява със стареенето, защото NRF2 предотвратява смъртта на стволовите клетки и им помага да се регенерират. NRF2 играе роля в подобряването на заздравяването на рани.
Укрепва съдовата система
Направено правилно с производството на сулфорафан, активирането на NRF2 може да предпази от сърдечни заболявания като високо кръвно налягане или хипертония и втвърдяване на артериите или атеросклероза. NRF2 може да засили релаксиращата активност на ацетилхолина или ACh върху съдовата система, като същевременно намалява стреса, предизвикан от холестерола. Активирането на Nrf2 може да укрепи сърцето, но свръхактивираният Nrf2 може да повиши вероятността от сърдечно-съдови заболявания.
Статините могат да предотвратят или да доведат до сърдечно-съдови заболявания. NRF2 също играе важна роля в балансирането на желязо и калций, които могат да предпазят човешкото тяло от повишени нива на желязо. Например, Sirtuin 2 или SIRT2 може да регулира хомеостазата на желязо в клетките чрез активиране на NRF2, който се смята, че е необходим за здрави нива на желязо. NRF2 може да помогне и при сърповидно-клетъчна болест или SCD. Дисфункцията на NRF2 може да бъде причина за ендотоксемия, като при дисбиоза или индуцирана от лектини хипертония. Nrf2 може също да предпази човешкото тяло от увреждане на съдовата система, предизвикано от амфетамин.
Бори се с невровъзпаление
NRF2 може да предпазва и подпомага възпалението на мозъка, обикновено наричано невровъзпаление. Освен това, NRF2 може да помогне при редица нарушения на централната нервна система или ЦНС, включително:
Болест на Алцхаймер (AD) – намалява амилоидния бета стрес върху митохондриите
Амиотрофична странична склероза (ALS)
болест на Хънтингтън (HD)
Множествена склероза (МС)
Регенерация на нервите
Болестта на Паркинсон (PD) – защитава допамина
Увреждане на гръбначния мозък (SCI)
Инсулт (исхемичен и хеморагичен) – подпомага хипоксията
Травматична мозъчна травма
NRF2 разкрива намаляване на невровъзпалението при тийнейджъри с разстройства от аутистичния спектър или ASD. Идебенон се сдвоява правилно с NRF2 активатори, за разлика от невровъзпалението. NRF2 може също да подобри кръвно-мозъчната бариера, или BBB. Например, активирането на NRF2 с карнозна киселина, получена от розмарин и градински чай, може да пресече BBB и да причини неврогенеза. Доказано е също, че NRF2 повишава мозъчния невротрофичен фактор или BDNF.
NRF2 също така модулира способността на някои хранителни добавки да предизвикат фактор на растежа на нервите или NGF, тъй като той също може да помогне при мозъчна мъгла и проблеми, предизвикани от глутамат, като модулира N-метил-D-аспартат или NMDA рецептори. Може също така да намали оксидативния стрес от хинолинова киселина, наричана QUIN. Активирането на NRF2 може да предпази от припадъци, а големи дози могат да намалят ръба на припадък. При редовни дози стимулация, NRF2 може да подобри когнитивните способности след припадък чрез понижаване на извънклетъчния глутамат в мозъка и чрез способността му да извлича цистеин от глутамат и глутатион.
Възстановява депресията
При депресия е нормално да забележите възпаление в мозъка, особено от префронталния кортекс и хипокампуса, както и намален BDNF. При някои версии на депресия NRF2 може да подобри симптомите на депресия чрез понижаване на възпалението в мозъка и повишаване на нивата на BDNF. Способността на Agmatine да намалява депресията чрез повишаване на норадреналин, допамин, серотонин и BDNF в хипокампуса зависи от активирането на NRF2.
Съдържа противоракови свойства
NRF2 е еднакво туморен супресор, както и туморен промотор, ако не се управлява съответно. NRF2 може да предпази от рак, причинен от свободни радикали и оксидативен стрес, но свръхекспресията на NRF2 може да бъде открита и в раковите клетки. Интензивното активиране на NRF2 може да помогне при различни видове рак. Например, добавката Protandim може да намали рака на кожата чрез активиране на NRF2.
Подобрява болката
Болест от войната в Персийския залив или GWI, забележително заболяване, засягащо ветераните от войната в Персийския залив, е съвкупност от необясними, хронични симптоми, които могат да включват умора, главоболие, болки в ставите, лошо храносмилане, безсъние, замаяност, респираторни заболявания и проблеми с паметта. NRF2 може да подобри симптомите на GWI чрез намаляване на хипокампалното и общото възпаление, в допълнение към намаляването на болката. NRF2 може допълнително да помогне при болка от увреждане на телесните нерви и да подобри увреждането на нервите от диабетна невропатия.
Подобрява диабета
Високите нива на глюкоза, най-добре наричани хипергликемия, причиняват окислително увреждане на клетките поради нарушаване на митохондриалната функция. Активирането на NRF2 може да защити човешкото тяло срещу увреждането на клетката от хипергликемия, като по този начин предотвратява клетъчната смърт. Активирането на NRF2 може допълнително да защити, възстанови и засили функцията на бета-клетките на панкреаса, като същевременно намали инсулиновата резистентност.
Защитава зрението и слуха
NRF2 може да предпази от увреждане на окото от диабетна ретинопатия. Може също така да избегне образуването на катаракта и да защити фоторецепторите срещу смърт, предизвикана от светлина. NRF2 допълнително предпазва ухото или кохлеята от стрес и загуба на слуха.
Може да помогне при затлъстяване
NRF2 може да помогне при затлъстяване главно поради способността си да регулира променливи, които действат върху натрупването на мазнини в човешкото тяло. Активирането на NRF2 със сулфорафан може да повиши инхибирането на синтеза на мастни киселини, или FAS, и разединяващите протеини, или UCP, което води до по-малко натрупване на мазнини и повече кафяви мазнини, характеризиращи се като мазнини, които включват повече митохондрии.
Предпазва червата
NRF2 помага за защита на червата, като защитава хомеостазата на чревния микробиом. Например пробиотиците от лактобацилус ще задействат NRF2, за да предпазят червата от оксидативен стрес. NRF2 може също да помогне за предотвратяване на улцерозен колит или UC.
Защитава половите органи
NRF2 може да защити тестисите и да предпази броя на сперматозоидите от увреждане при хора с диабет. Може също да помогне при еректилна дисфункция или ЕД. Някои добавки за повишаване на либидото, като Mucuna, Tribulus и Ashwaganda, могат да подобрят сексуалната функция чрез активиране на NRF2. Други фактори, които повишават NRF2, като слънчева светлина или кълнове от броколи, също могат да помогнат за подобряване на либидото.
Регулира костите и мускулите
Оксидативният стрес може да доведе до намаляване на костната плътност и сила, което е нормално при остеопороза. Активирането на NRF2 може да има способността да подобри антиоксидантите в костите и да предпази от стареене на костите. NRF2 може също да предотврати загубата на мускули и да засили мускулната дистрофия на Дюшен или DMD.
Съдържа антивирусни свойства
Не на последно място, активирането на NRF2 може в крайна сметка да помогне за защитата на човешкото тяло срещу няколко вируса. При пациенти с вируса на денга симптомите не са били толкова интензивни при индивиди, които са имали по-високи нива на NRF2 в сравнение с индивиди, които са имали по-ниски степени на NRF2. NRF2 може също да помогне на хора, които имат вирус на човешки имунодефицит-1 или ХИВ. NRF2 може да предпази от оксидативния стрес от адено-асоциирания вирус, или AAV, и H. Pylori. И накрая, Lindera Root може да потисне вируса на хепатит С с активиране на NRF2.
Nrf2, или NF-E2-свързан фактор 2, е транскрипционен фактор, открит при хора, който регулира експресията на специфичен набор от антиоксидантни и детоксикиращи гени. Този сигнален път се активира поради оксидативния стрес, тъй като засилва множество антиоксидантни и ензими за детоксикация на черния дроб фаза II за възстановяване на хомеостазата в човешкото тяло. Хората са адаптирани да функционират в състояние на хомеостаза или баланс. Когато тялото е изправено пред оксидативен стрес, Nrf2 се активира, за да регулира окисляването и да контролира стреса, който причинява. Nrf2 е от съществено значение за предотвратяване на здравословни проблеми, свързани с оксидативния стрес. Д-р Алекс Хименес DC, CCST Insight
Сулфорафан и неговите ефекти върху рака, смъртността, стареенето, мозъка и поведението, сърдечните заболявания и др.
Изотиоцианатите са едни от най-важните растителни съединения, които можете да получите във вашата диета. В това видео правя най-изчерпателния случай за тях, който някога е бил направен. Кратко внимание? Прескочи до любимата си тема, като кликнете върху някоя от точките по-долу. Пълна времева линия по-долу.
Ключови секции:
00: 01: 14 - Рак и смъртност
00: 19: 04 - Стареене
00: 26: 30 - Мозък и поведение
00: 38: 06 - Краен рекап
00: 40: 27 - Доза
Пълна времева линия:
00: 00: 34 - Въвеждане на сулфорафан, основен фокус на видеото.
00: 01: 14 - Консумация на кръстоцветни зеленчуци и намаляване на смъртността от всякаква причина.
00: 02: 12 - риск от рак на простатата.
00: 02: 23 - риск от рак на пикочния мехур.
00: 02: 34 - Рак на белия дроб при риск от пушене.
00: 02: 48 - риск от рак на гърдата.
00: 03: 13 - Хипотетично: какво ще стане, ако вече имате рак? (Интервенция)
00: 03: 35 - Вероятен механизъм, който управлява асоциативните данни за рака и смъртността.
00: 04: 38 - Сулфорафан и рак.
00: 05: 32 - доказателства при животни, показващи силен ефект на екстракт от кълнове от броколи върху развитието на тумор на пикочния мехур при плъхове.
00: 06: 06 - Ефект на директно добавяне на сулфорафан при пациенти с рак на простатата.
00: 07: 09 - Биоакумулиране на изотиоцианатни метаболити в действителната гръдна тъкан.
00: 08: 32 - Инхибиране на стволови клетки от рак на гърдата.
00: 08: 53 - История: Brassicas са установени със здравни качества дори в древния Рим.
00: 09: 16 - Способността на Sulforaphane да подобри екскрецията на канцероген (бензен, акролеин).
00: 09: 51 - NRF2 като генетичен превключвател чрез елементи на антиоксидантен отговор.
00: 10: 10 - Как активирането на NRF2 подобрява екскрецията на канцероген чрез глутатион-S-конюгати.
00: 10: 34 - Брюкселското зеле увеличава глутатион-8-трансферазата и намалява увреждането на ДНК.
00: 11: 20 - Напитката от кълнове от броколи повишава екскрецията на бензен с 61%.
00: 15: 45 - Кръстосана консумация на зеленчуци и смъртност от сърдечни заболявания.
00: 16: 55 - Броколи зеленчуков прах подобрява кръвните липиди и цялостния риск от сърдечни заболявания при диабетици тип 2.
00: 19: 04 - начало на стареене секция.
00: 19: 21 - Диета, обогатена със сулфарафан, увеличава живота на бръмбарите от 15 до 30% (при определени условия).
00: 20: 34 - Значение на ниското възпаление за дълголетие.
00: 22: 05 - Кръстоцветните зеленчуци и кълнове от кълнове от броколи изглежда намаляват голямо разнообразие от възпалителни маркери при хората.
00: 23: 40 - Средно видеоклип: рак, стареене
00: 24: 14 - Изследванията на мишки показват, че сулфарафан може да подобри адаптивната имунна функция в напреднала възраст.
00: 25: 18 - Сулфарафан подобри растежа на косата при миши модел на оплешивяване. Изображение на 00: 26: 10.
00: 26: 30 - начало на секцията за мозъка и поведението.
00: 27: 18 - Ефект на екстракт от кълнове от броколи върху аутизма.
00: 27: 48 - Ефект на глюкорафанин върху шизофрения.
00: 28: 17 - Начало на депресивната дискусия (правдоподобен механизъм и проучвания).
00: 31: 21 - Изследване на мишки, използващи различни модели 10 на стресова индуцирана депресия, показва сулфорафан с подобен ефект като флуоксетин (prozac).
00: 32: 00 - Изследването показва, че директното поглъщане на глюкорафанин в мишки е подобно ефективно при предотвратяване на депресията от модела на социално потискане на стреса.
00: 33: 01 - начало на секцията за невродегенерация.
00: 33: 30 - Сулфорафан и болестта на Алцхаймер.
00: 33: 44 - Сулфорафан и болестта на Паркинсон.
00: 33: 51 - Сулфорафан и болестта на Хънтингтън.
00: 34: 13 - Сулфорафан увеличава протеините на топлинния шок.
00: 34: 43 - Начало на травматичното мозъчно увреждане.
00: 35: 01 - Сулфарафан се инжектира веднага след като TBI подобрява паметта (изследване на мишката).
00: 35: 55 - Сулфорафан и невронна пластичност.
00: 36: 32 - Сулфорафан подобрява ученето при модел на диабет тип II при мишки.
00: 37: 19 - Сулфорафан и душевна мускулна дистрофия.
00: 37: 44 - Инхибиране на миостатина в мускулните сателитни клетки (in vitro).
00: 38: 06 - Късно видеоклип: смъртност и рак, увреждане на ДНК, оксидативен стрес и възпаление, екскреция на бензол, сърдечно-съдови заболявания, диабет тип II, ефекти върху мозъка (депресия, аутизъм, шизофрения, невродегенерация).
00: 40: 27 - Мисли за определяне на доза от кълнове от броколи или сулфорафан.
00: 41: 01 - Анекдоти за поникване у дома.
00: 43: 14 - На температурата на готвене и активността на сулфорафан.
00: 43: 45 - Конверсия на сулфорафан от глюкорафанин в чревната бактерия.
00: 44: 24 - Добавките работят по-добре, когато се комбинират с активна мирозиназа от зеленчуци.
00: 44: 56 - техники за готвене и кръстоцветни зеленчуци.
00: 46: 06 - Изотиоцианати като глюкогени.
Когато човешкото тяло е изправено пред вредни вътрешни и външни фактори като токсини, клетките трябва бързо да задействат своите антиоксидантни способности, за да противодействат на оксидативния стрес. Тъй като е установено, че повишените нива на оксидативен стрес причиняват различни здравословни проблеми, важно е да използвате Nrf2 активиране, за да се възползвате от неговите предимства. Обхватът на нашата информация е ограничен до хиропрактика и проблеми със здравето на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Допълнителна дискусия на тема: „Остра болка в гърба
Заболявания на опорно-двигателния апаратЕ една от най-разпространените причини за увреждане и пропуснати дни на работа в световен мащаб. Болките в гърба се дължат на втората най-честа причина за посещения в лекарски кабинет, превъзхождани само от инфекции на горните дихателни пътища. Приблизително 80% от населението ще изпитва болки в гърба поне веднъж през живота си. Гръбначният стълб е сложна структура, съставена от кости, стави, връзки и мускули, наред с други меки тъкани. Поради това наранявания и / или влошени състояния катохерния дискове, в крайна сметка може да доведе до симптоми на болки в гърба. Спортните наранявания или нараняванията при автомобилни катастрофи често са най-честата причина за болки в гърба, но понякога най-простите движения могат да имат болезнени резултати. За щастие, алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Sulforaphane е фитохимикал, вещество от изотиоцианатната група на органосерни съединения, намиращи се в кръстоцветни зеленчуци, като броколи, зеле, карфиол и брюкселско зеле. Може да се намери и в бок чой, кейл, зеле, горчица и кресон. Изследванията показват, че сулфорафанът може да помогне за предотвратяване на различни видове рак чрез активиране на производството на Nrf2, или ядрен фактор, свързан с еритроид 2 фактор, транскрипционен фактор, който регулира защитните антиоксидантни механизми, които контролират реакцията на клетката към оксиданти. Целта на следващата статия е да опише функцията на сулфорафан.
абстрактен
Антиоксидантната система KEAP1-Nrf2-ARE е основно средство, чрез което клетките реагират на оксидативния и ксенобиотичния стрес. Сулфорафанът (SFN), електрофилен изотиоцианат, получен от кръстоцветни зеленчуци, активира пътя KEAP1-Nrf2-ARE и се превърна в молекула от интерес при лечението на заболявания, при които хроничният оксидативен стрес играе основна етиологична роля. Тук демонстрираме, че митохондриите на култивирани клетки от човешки пигментен епител на ретината (RPE-1), третирани със SFN, претърпяват хиперфузия, която е независима както от Nrf2, така и от неговия цитоплазмен инхибитор KEAP1. Съобщава се, че митохондриалното сливане е цитопротективно чрез инхибиране на образуването на пори в митохондриите по време на апоптоза и в съответствие с това, ние показваме Nrf2-независима, цитопротекция на SFN-третирани клетки, изложени на индуктор на апоптоза, ставроспорин. Механично, SFN смекчава набирането и/или задържането на разтворимия фактор на делене Drp1 към митохондриите и пероксизомите, но не влияе върху общото изобилие на Drp1. Тези данни показват, че полезните свойства на SFN се простират отвъд активирането на системата KEAP1-Nrf2-ARE и изискват по-нататъшен разпит, като се има предвид текущата употреба на този агент в множество клинични проучвания.
Сулфорафанът е Nrf2-независим инхибитор на митохондриалното делене
Сулфорафанът (SFN) е изотиоцианатно съединение, получено в диетата най-често от кръстоцветни зеленчуци [56]. Той се генерира в растенията като ксенобиотичен отговор на хищничество чрез везикуларно освобождаване на хидролитичния ензим мирозиназа от увредените клетки; този ензим превръща глюкозинолатите в изотиоцианти [42]. През последните две десетилетия SFN е широко характеризиран със своите докладвани противоракови, антиоксидантни и антимикробни свойства [57]. Голяма част от тази ефикасност се дължи на способността на SFN да модулира сигналния път на KEAP1-Nrf2-антиоксидантния отговорен елемент (ARE), въпреки че са идентифицирани допълнителни дейности на съединението, включително инхибиране на активността на хистон деацетилаза и прогресия на клетъчния цикъл [ 29]. Nrf2 е главният антиоксидантен транскрипционен фактор и при условия на хомеостаза неговата стабилност се потиска чрез действието на цитоплазмения Cullin3KEAP1 убиквитин лигазен комплекс [20]. По-конкретно, Nrf2 се набира към лигазата Cullin3KEAP1 чрез свързване към димерния субстратен адаптер KEAP1 и впоследствие се модифицира с polyUb вериги, които са насочени към транскрипционния фактор за протеазомно-медиирано разграждане. Този конститутивен оборот ограничава полуживота на Nrf2 в нестресирани клетки до ~ 15 минути [30], [33], [46], [55]. В отговор на множество видове стрес, най-вече оксидативен стрес, KEAP1, богат на цистеин протеин, действа като редокс сензор, а окислителното модифициране на критични цистеини, особено C151, на KEAP1 дисоциира Nrf2-KEAP1 от CUL3, като по този начин предотвратява разграждането на Nrf2 [ 8], [20], [55]. По-специално, SFN и вероятно други Nrf2 активатори, имитират оксидативен стрес чрез модифициране на C151 на KEAP1, например [21]. Стабилизирането на Nrf2 позволява неговата транслокация към ядрото, където индуцира експресията на батерия от фаза II антиоксидантни и детоксикационни гени. Nrf2 се свързва с промоторните елементи на антиоксидантния отговор (ARE) на неговите сродни целеви гени чрез хетеродимеризация с малки Maf протеини [19]. Тази система представя динамичен и чувствителен отговор към индиректни антиоксиданти като SFN, свободни радикали, генерирани от митохондриите [16], или други физиологични източници на оксидативен стрес [41].
Митохондриите са динамични субклетъчни органели, които регулират множество клетъчни функции, вариращи от производство на АТФ и вътреклетъчно калциево буфериране до редокс регулация и апоптоза [13], [49]. Митохондриите също представляват основният източник на реактивни кислородни видове (ROS) в клетката. Следователно е необходимо правилно регулиране на митохондриалната функция за оптимизиране на производството на АТФ, за да отговори на клетъчните нужди, като същевременно се минимизират потенциално вредните ефекти от прекомерното производство на свободни радикали. Критично изискване за фина модулация на митохондриалната функция е способността на митохондриите да функционират както независимо като биохимични машини, така и като част от обширна, отзивчива мрежа.
Морфологията и функцията на митохондриалната мрежа се определят от регулиран баланс между делене и сливане. Митохондриалното делене е необходимо за наследяване на дъщерната клетка на митохондриите по време на клетъчното делене [28], както и за селективното, автофагично разграждане на деполяризирани или увредени митохондрии, наречено митофагия [1]. Обратно, сливането е необходимо за допълване на митохондриалните геноми и споделяне на компоненти на веригата за транспорт на електрон между съседни митохондрии [54]. На молекулярно ниво, деленето и сливането на митохондриите се регулират от големи, подобни на динамин GTPази. Три ензима основно регулират сливането: Митофузините 1 и 2 (Mfn1/2) са двупроходни външни мембранни протеини, които медиират сливането на външната мембрана чрез хетеротипни взаимодействия между съседни митохондрии [15], [25], [37], докато OPA1 е вътрешен мембранен протеин, който едновременно осигурява свързване на матрицата чрез регулиране на сливането на вътрешните мембрани [5]. Активността на GTPase и на трите протеина е необходима за стабилно сливане [5], [18], а OPA1 се регулира допълнително чрез сложна протеолиза в митохондриалната вътрешна мембрана от протеазите OMA1 [14], PARL [6] и YME1L [45 ]. Важно е, че потенциалът на непокътнатата митохондриална мембрана е необходим за ефективно сливане, за да се потисне интеграцията на увредени и здрави митохондрии [26].
Митохондриалното делене се катализира предимно от цитозолен протеин, наречен Dynamin-свързан протеин 1 (Drp1/DNM1L). Drp1 се набира от цитозола до проспективни места на делене на външната мембрана на митохондриите [43]. Основните рецептори за Drp1 на външната мембрана са митохондриалният фактор на делене (Mff) [32] и в по-малка степен Fission 1 (Fis1) [51]. Освен това е открит рецептор за примамка, MIEF1/MiD51, който действа за допълнително ограничаване на активността на протеина Drp1 в потенциални места на делене [58]. Веднъж прикрепен към външната мембрана на митохондриите, Drp1 олигомеризира в спираловидни структури около тялото на митохондриона и след това използва енергията, получена от хидролизата на GTP, за да посредничи при физическото разрязване на външните и вътрешните мембрани на митохондриите [17]. Тубули, получени от ендоплазмен ретикулум, действат като начален констриктор на митохондриите преди олигомеризацията на Drp1, подчертавайки разкритието, че неконстрикираните митохондрии са по-широки от допустимата обиколка на завършена Drp1 спирала [12]. Динамиката на актина също е важна за взаимодействията на ER-митохондриите, които предхождат митохондриалното делене [24]. В допълнение към ролята си в деленето на митохондриите, Drp1 катализира деленето на пероксизомите [40].
Drp1 е много подобен на добре характеризирания динамин протеин по това, че и двата протеина съдържат N-терминален GTPase домен, среден домен, който е критичен за самоолигомеризация, и C-терминален GTPase ефекторен домен [31]. Drp1 постига селективност за митохондриалните мембрани чрез комбинация от взаимодействия с неговите рецепторни протеини Mff и Fis1, както и чрез неговия афинитет към специфичния за митохондриите фосфолипиден кардиолипин чрез уникалния B-вмъкващ домейн на Drp1 [2]. Drp1 обикновено съществува като хомотетрамер в цитоплазмата, а сглобяването от по-висок порядък в местата на митохондриално делене се медиира от средния домейн на Drp1 [3].
Като се има предвид имплицитната връзка между митохондриалната функция и пътя KEAP1-Nrf2-ARE, ние изследвахме ефектите от активирането на Nrf2 върху митохондриалната структура и функция. Тук демонстрираме, че SFN индуцира митохондриална хиперфузия, която неочаквано е независима както от Nrf2, така и от KEAP1. Този ефект на SFN е чрез инхибиране на функцията Drp1. Освен това демонстрираме, че SFN придава резистентност към апоптоза, която е Nrf2-независима и имитира наблюдаваната в клетки, изчерпани от Drp1. Тези данни заедно показват, че в допълнение към стабилизирането и активиране на Nrf2, SFN модулира митохондриалната динамика и запазва клетъчната годност и оцеляване.
Резултати
Сулфорафанът предизвиква Nrf2/KEAP1-независима хиперфузия на митохондриите
В хода на изучаването на ефектите от активирането на Nrf2 върху динамиката на митохондриалната мрежа, ние открихме, че лечението на обезсмъртени клетки на човешки пигментен епител на ретината (RPE-1) със сулфорафан (SFN), мощен активатор на Nrf2 сигнализация, предизвиква стабилно сливане на митохондриалната мрежа в сравнение с контролни клетки, третирани с носител (фиг. 1А и Б). Морфологията на митохондриите в тези клетки много наподобява тази на митохондриите в клетки, изчерпани от siRNA на ендогенния Drp1, основният фактор на митохондриално делене (Фиг. 1А). Този резултат повдигна интригуващата идея, че състоянието на митохондриално делене и сливане реагира директно на нивата на Nrf2 в клетката. Въпреки това, стимулирането на клетките с други Nrf2 стабилизатори и активатори, като протеазомния инхибитор MG132, прооксидантът tBHQ, или нокдаун на Nrf2 инхибитора KEAP1 не индуцира митохондриално сливане (фиг. 1А и В). Стабилизирането на Nrf2 чрез тези манипулации беше потвърдено чрез Western blotting за ендогенен Nrf2 (фиг. 1C). Освен това, експресията на Nrf2 беше незаменима за индуцирано от SFN митохондриално сливане, тъй като нокдаунът на ендогенен Nrf2 с siRNA не успя да противодейства на този фенотип (фиг. 1D F). Тъй като SFN стимулира пътя KEAP1-Nrf2-ARE чрез ковалентно модифициране на цистеинови остатъци на KEAP1 [21], ние съборихме KEAP1, за да разгледаме дали индуцираната от SFN митохондриална хиперфузия се стимулира чрез KEAP1-зависим, но независим от Nrf2 път. Въпреки това, изчерпването на KEAP1 също не успя да отмени индуцираното от SFN митохондриално сливане (фиг. 1G I). Всъщност, SFN обърна морфологията на професията, предизвикана от изчерпването на KEAP1 (фиг. 1G, панел b срещу панел d). Тези резултати показват, че лечението с SFN причинява митохондриално сливане, независимо от каноничния път KEAP1-Nrf2-ARE и ни накара да се запитаме дали SFN пряко засяга компонентите на митохондриалното делене или машината за синтез.
Фигура 1 SFN индуцира Nrf2/KEAP1-независимо митохондриално сливане. (A) RPE-1 клетките бяха трансфектирани с посочените siRNAs и 3 дни по-късно бяха третирани с DMSO или Nrf2 активаторите SFN (50 цМ), MG132 (10 цМ) или tBHQ (100 цМ) за 4 часа. Митохондриите (червени) са белязани с анти-Tom20 антитяло, а ядрата (синьо) са противооцветени с DAPI. (B) Графика, показваща количествена оценка на митохондриалната морфология от (A). >50 клетки на състояние бяха оценени по заслепен начин. (C) Представителни уестърн блотове от (A). (D) RPE-1 клетките бяха трансфектирани с 10 nM siRNA и 3 дни по-късно бяха третирани със SFN в продължение на 4 часа, преди да бъдат фиксирани и оцветени, както в (А). (E) Графика, показваща количествено определяне на оценката на митохондриалния фенотип от (D). >100 клетки на състояние бяха оценени по заслепен начин. (F) Представителни уестърн блотове от (D). (G) Клетките бяха трансфектирани и третирани както в (D) със siCON или siKEAP1. (H) Клетките от (G) се оценяват както в (B) и (E) въз основа на митохондриална морфология. (I) Представителни уестърн блотове от (G). Данните в (B), (E) и (H) бяха събрани от 3 независими експеримента всеки и статистическата значимост беше определена чрез двустранния t-тест на Student. Лентите за грешки отразяват +/- SD (За тълкуване на препратките към цвета в тази легенда на фигурата, читателят се препраща към уеб версията на тази статия).
Сулфорафанът уврежда митохондриалната асоциация на Drp1
Въз основа на констатацията, че лечението със SFN индуцира митохондриална хиперфузия, ние аргументирахме, че този фенотип е или следствие от прекомерна фузионна активност, или инхибиране на активността на делене. За да разграничим тези две възможности, ние сравнихме морфологията на пероксизомите в присъствието и отсъствието на SFN. Пероксизомите са подобни на митохондриите по това, че са динамични органели, чиято форма и дължина са постоянно в поток [44]. Пероксизомите съдържат както Fis1, така и Mff във външната си мембрана и в резултат на това са мишени за Drp1-медиирано делене [22], [23]. Въпреки това, пероксизомите не използват механизмите за сливане на митохондриалната мрежа и следователно не се подлагат на сливане [39]. По-скоро, пероксизомното делене се противопоставя от удължаването на съществуващите пероксизоми чрез de novo добавяне на мембрани и протеини [44]. Тъй като в пероксизомите липсват Mfn1/2 и OPA1, ние разсъждавахме, че ако SFN активира машината за сливане, а не инхибира машината за делене, дължината на пероксизомата няма да бъде засегната. В клетки, третирани с носител, пероксизомите се поддържат като къси, кръгли, точковидни органели (фиг. 2, панели b и d). Въпреки това, лечението със SFN увеличава дължината на пероксизома с ~ 2 пъти в сравнение с контролните клетки (фиг. 2, панели f и h). Освен това, много от пероксизомите бяха прищипани близо до центъра, което показва потенциален дефект на разкъсване (фиг. 2, панел h, върхове на стрелки). По същия начин, пероксизомите в клетки, трансфектирани с Drp1 siRNA, бяха необичайно дълги (фиг. 2, панели j и l), потвърждавайки, че Drp1 е необходим за пероксизомно делене и предполага, че SFN-лечение причинява митохондриални и пероксизомни фенотипове чрез разрушаване на машината за делене.
Фигура 2 SFN индуцира пероксизомно удължаване. (A) RPE-1 клетките бяха трансфектирани с 10 nM от посочената siRNA и 3 дни по-късно бяха третирани с DMSO или 50 цМ SFN за 4 часа. Пероксизомите (зелени) бяха белязани с анти-PMP70 антитяло, митохондриите с MitoTracker (червени) и ДНК, противооцветена с DAPI. Отдясно са показани увеличени вмъквания на пероксизоми (панели d, h и l), за да се улесни визуализацията на промените в морфологията, предизвикани от изчерпването на SFN и Drp1. Стрелките подчертават точките на стесняване. (За тълкуване на препратките към цвета в тази легенда на фигурата, читателят се препраща към уеб версията на тази статия).
След това определихме как SFN ограничава функцията Drp1. Възможностите включват намаляване на нивата на експресия, набиране/задържане в митохондриите, олигомеризация или ензимна активност на GTPase. Дефицитът на някое от тях би довел до намалено делене на митохондриите и хиперфузия. Ние не открихме възпроизводими промени в нивата на протеин Drp1 след лечение със SFN (фиг. 1C и 3A) и следователно заключихме, че SFN не променя стабилността или експресията на Drp1, в съответствие с Drp1 с полуживот >10 часа [50] и нашите SFN лечения са с по-кратка продължителност. След това проучихме дали SFN повлиява набирането или задържането на Drp1 в митохондриите. Проучванията за фракциониране показват, че SFN индуцира загуба на Drp1 от митохондриалната фракция (фиг. 3А, ленти 7 8 и фиг. 3В). Както беше съобщено по-рано [43], само малка част от Drp1 (~3%) е свързана с митохондриалната мрежа по всяко време по време на стационарни условия с по-голямата част от ензима, намиращ се в цитоплазмата (Фиг. 3А, ленти 5 8 ). Тези данни за фракциониране бяха потвърдени с помощта на анализ на съвместна локализация, който показа ~40% намаление на локализираните в митохондриите, точковидни Drp1 фокуси след SFN-лечение (фиг. 3C и D). Заедно тези данни показват, че митохондриалното сливане, индуцирано от SFN, се дължи поне частично на отслабената връзка на Drp1 с митохондриите. Нашите данни не правят разлика между това дали SFN пречи на набирането на митохондриите спрямо митохондриалното задържане на Drp1, или и двете, тъй като анализът на ендогенния Drp1 не е податлив на визуализиране на GTPase чрез микроскопия на живи клетки.
Фигура 3 SFN причинява загуба на Drp1 от митохондриите. (A) Субклетъчно фракциониране на RPE-1 клетки след 4 часа DMSO или SFN. Цялоклетъчни лизати (WCL), ядрени (Nuc), цитозолни (Cyto) и сурови митохондриални (Mito) фракции се разделят чрез SDS-PAGE и се обработват за Western blotting с посочените антитела. Миграцията на маркерите за молекулно тегло е посочена вляво. (B) Графики, показващи денситометрично количествено определяне на Drp1 в посочените фракции от (A). (C) RPE-1 клетките бяха трансфектирани с 10 nM siCON или siDrp1 и 3 дни по-късно бяха третирани с DMSO или SFN в продължение на 4 часа. Drp1 (зелен) се визуализира с анти-Drp1 антитяло, митохондриите с MitoTracker (червено) и ядрата с DAPI (синьо). (D) Автоматичен анализ на съвместната локализация на сигнала Drp1 и MitoTracker от (C). Данните в (B) и (D) са събрани от 3 и 5 независими експеримента, съответно, и статистическата значимост е определена чрез двустранния t-тест на Student. Лентите за грешки отразяват +/- SD, а звездичките означават статистическа значимост. (За тълкуване на препратките към цвета в тази легенда на фигурата, читателят се препраща към уеб версията на тази статия).
Сулфорафанът осигурява защита срещу индуцирана от стауроспортин апоптоза, независима от Nrf2
Предишна работа показа, че митохондриалното делене е разрешително при образуването на пори във външната митохондриална мембрана, генерирани от Bax/Bak по време на апоптоза [11]. Доказано е, че Drp1 се набира селективно в митохондриите по време на апоптоза [11] и, в съответствие с това, фрагментирани митохондрии са наблюдавани в началото на процеса [27]. Обратно, смята се, че инхибирането на митохондриалното делене инхибира апоптозата чрез блокиране на образуването на пори на външната мембрана, които позволяват освобождаване на цитохром с [53]. Съответно, стимулирането на митохондриално сливане забавя прогресията на апоптозата, индуцирана от съединения, включително ставроспорин (STS) [14]. За да определим дали SFN защитава RPE-1 клетки от STS-медиирана апоптоза и ако е така, дали това изисква Nrf2, ние създадохме анализ за лесно индуциране на разцепване на поли ADP рибоза полимераза (PARP), субстрат на активирана каспаза-3 и окончателен маркер на апоптоза. Третирането на RPE-1 клетки с 1 µM STS в продължение на 6 часа причинява само много скромно разцепване на PARP, но това е предотвратено чрез SFN съвместно третиране (например, Фиг. 4А, лента 3 срещу 4). За да увеличим устойчивостта на този анализ, ние допълнително сенсибилизирахме клетките към индуцирана от STS апоптоза, като ги третирахме предварително със siRNA, насочена към антиапоптотичния фактор, Bcl-XL. Тази предварителна обработка намалява експресията на Bcl-XL и значително насърчава разцепването на PARP като функция от времето, изложено на STS (фиг. 4В, сравнете лента 2 с ленти 4 10). Важно е, че 2 часа предварително третиране със SFN смекчава разцепването на PARP в клетки, изложени на STS (фиг. 4C, лента 3 срещу 4 и лента 5 срещу 6). По същия начин, клетките, стабилно изчерпани от Nrf2 от CRISPR/Cas9, бяха сравнително защитени от STS токсичност чрез SFN предварителна обработка (фиг. 4C, лента 11 срещу 12 и лента 13 срещу 14 и фиг. 4D). Тази защита се наблюдава при използване както на PARP разцепване (фиг. 4C и D), така и на клетъчна морфология (фиг. 4E) като показания. Ефикасността на изчерпването на Nrf2 от CRISPR/Cas9 беше потвърдена чрез Western blotting (фиг. 4C, Nrf2 blot). Както беше предвидено, изчерпването на клетките на Drp1, което също води до хиперфузионен фенотип (фиг. 1А), също блокира разцепването на PARP в отговор на STS в сравнение с контролните клетки, инкубирани със SFN (фиг. 4F и G). Заедно тези констатации са в съответствие с SFN, предоставящ защита срещу апоптоза чрез способността си да ограничава функцията на Drp1, независимо от стабилизирането и активирането на Nrf2.
Фигура 4 Цитопротективните ефекти на SFN са независими от експресията на Nrf2 (A) RPE-1 клетките бяха предварително третирани с DMSO или 50 цМ SFN в продължение на 2 часа преди третиране с DMSO, 1 цМ ставроспорин (STS) или 50 цМ SFN. М етопозид за 6 часа и се обработват за анти-PARP Western blotting. (B) RPE-1 клетките бяха трансфектирани с 2.5 nM siCON, 1 nM siBcl-XL или 2.5 nM siBcl-XL и 3 дни по-късно бяха третирани с DMSO или 1 цМ STS за 2, 4 или 6 часа. Показани са представителни Western блотове и миграцията на маркери за молекулно тегло е посочена вляво. (C) CRISPR/Cas9-генериран див тип (Nrf2WT) и Nrf2 нокаут (Nrf2KO) RPE-1 клетки бяха трансфектирани с 1 nM siBcl-XL и 3 дни по-късно бяха предварително третирани с DMSO или 50 цМ SFN за 2 часа . След това клетките се третират с 1 цМ STS за 2, 4 или 6 часа. Показани са представителни Western блотове с посочените антитела. (D) Количествено определяне на разцепения PARP като процент от общия PARP (разцепен+неразцепен) от 3 независими експеримента. Важно е, че нивата на разцепен PARP са сравними независимо дали клетките експресират Nrf2 или не, което показва, че SFN защитата от STS е независима от транскрипционния фактор. (E) 20X фазово-контрастни изображения, направени непосредствено преди събирането на лизати от (C). Мащабна лента=65 m. (F) Представителни Western блотове, демонстриращи, че изчерпването на Drp1 дава почти сравнима защита от STS като SFN лечение. RPE-1 клетките бяха трансфектирани с 1 nM siBcl-XL и допълнително трансфектирани с 10 nM siCON или 10 nM siDrp1. 3 дни по-късно, siCON клетките бяха предварително третирани със SFN, както в (A) и (C) и след това изложени на STS за 4 часа преди да бъдат събрани и обработени за Western blotting с посочените антитела. (G) Същото като (D) за данните, представени в (F), събрани от 3 независими експеримента. Лентите за грешки отразяват +/- SEM
Дискусия
Ние открихме, че SFN модулира динамиката на митохондриално делене/сливане независимо от ефектите му върху пътя KEAP1-Nrf2-ARE. Това е интригуващо поради предполагаемата връзка между митохондриалната дисфункция и производството на ROS и необходимостта от потискане на свободните радикали, получени от митохондриите, чрез активиране на Nrf2. Това допълнително функционално въздействие на SFN е от потенциално значение, като се имат предвид над 30 клинични проучвания, които в момента се провеждат, тестващи SFN за лечение на различни заболявания, включително рак на простатата, обструктивна белодробна болест и сърповидно-клетъчна болест [7], [10], [ 47].
Тъй като SFN е изотиоцианат [56] и активира Nrf2 сигнализиране чрез директно ацилиране на критичните KEAP1 цистеини за потискане на разграждането на Nrf2 [21], следва, че SFN упражнява своите про-фузионни ефекти чрез модулиране на активността на делене или сливане на фактор чрез модификация на цистеин . Нашите данни силно подкрепят, че Drp1 се регулира отрицателно от SFN, въпреки че остава да се изясни дали GTPase е пряка цел на ацилиране. Въпреки тази празнина в знанията, функцията на Drp1 очевидно е компрометирана от SFN, тъй като и митохондриите, и пероксизомите стават хиперфузирани в отговор на лечението със SFN и тези органели споделят Drp1 за съответните им събития на разкъсване [38]. В допълнение, SFN намалява количеството Drp1, което се локализира и натрупва в митохондриите (фиг. 3). Тъй като нашите експерименти бяха направени с всички ендогенни протеини, нашето откриване на Drp1 в местата на митохондриално делене е при стационарни условия и следователно не можем да направим разлика между набиране и дефект на задържане на ензима, причинен от SFN. Освен това, не можем да елиминираме възможността SFN да ацилира рецептор в митохондриите (Fis1 или Mff), за да блокира набирането на Drp1, все още подозираме, че Drp1 е директно модифициран. Drp1 има девет цистеина, осем от които се намират в средния домейн, който е необходим за олигомеризация [3], и един от които се намира в GTPase Effector Domain (GED) в C-края на Drp1. Директното ацилиране на всеки от тези цистеини може да причини дефект на активността в Drp1 и следователно да е в основата на ефекта на SFN върху митохондриалната динамика. По-специално, предишната работа предполага, че дефектите в олигомеризацията и каталитичната активност могат да отменят задържането на Drp1 в митохондриите [52]. Cys644 в GED домейна е особено атрактивна цел въз основа на предишна работа, показваща, че мутацията на този цистеин фенокопира мутации, които нарушават активността на Drp1 GTPase [4] и че този конкретен цистеин е модифициран от тиол-реактивни електрофили [9]. Решаването на този нерешен въпрос ще изисква масспектрометрично валидиране. В обобщение, ние идентифицирахме нова, цитопротективна функция за клинично значимото съединение SFN. В допълнение към активиране на главния антиоксидантен транскрипционен фактор Nrf2, SFN насърчава митохондриалното и пероксизомното сливане и този ефект е независим от Nrf2. Механизмът, лежащ в основата на това явление, включва намаляване на функцията на GTPase Drp1, основният медиатор на митохондриалното и пероксизомното делене. Основна последица от SFN-медиирано митохондриално сливане е, че клетките стават резистентни към токсичните ефекти на индуктора на апоптоза стауроспорин. Това допълнително цитопротективно действие на SFN може да бъде от особена клинична полза при множество невродегенеративни заболявания, за които възрастта е водещ рисков фактор (напр. болест на Паркинсон, болест на Алцхаймер, свързана с възрастта макулна дегенерация), тъй като тези заболявания са свързани с апоптоза и намаляват нива и/или дисрегулация на Nrf2 [35], [36], [48].
Материали и методи
Анализи на апоптоза
Клетките се посяват и трансфектират с siRNA, както е посочено по-долу. Клетките бяха предварително третирани с 50 цМ сулфорафан в продължение на 2 часа за индуциране на митохондриално сливане и след това бяха третирани с 1 цМ ставроспорин за индуциране на апоптоза. По време на прибиране на реколтата средата се събира в отделни епруветки и се подлага на високоскоростно центрофугиране до пелетни апоптотични клетки. Тази клетъчна пелета се комбинира с прилепнали клетки и се разтваря в 2 пъти концентриран Laemmli буфер. Пробите бяха подложени на анти-PARP Western blotting.
Генериране на конструкция CRISPR/Cas9
За да създадете LentiCRISPR/eCas9 1.1, LentiCRISPR v2 (addgene #52961) първо беше изрязан с Age1 и BamH1. След това SpCas9 от eSpCas9 1.1 (addgene #71814) беше PCR амплифициран с Age1 и BamH1 надвеси, като се използват следните праймери (Напред AGCGCACCGGTTCTAGAGCGCTGCCACCATGGACTATAAGGACCACGAC, Обратно AAGCGCGGATCCCCTTCGTTGTTGTTGTTGTTGTTGTTGTTGTTGTTGTTGTTGTTGTTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTG sgRNA последователностите бяха определени чрез използване на Benchling.com. Параметрите бяха зададени така, че да насочват кодиращата последователност с най-високи резултати при прицел и най-ниски извън целта. Следващите последователности (насочване последователност подчертани, ХС sgNFE2L2 # 1 смисъл CACCGCGACGGAAAGAGTATGAGC, антисенс AAACGCTCATACTCTTTCCGTCGC; HS sgNFE2L2 # 2 смисъл CACCGGTTTCTGACTGGATGTGCT, антисенс AAACAGCACATCCAGTCAGAAACC; HS sgNFE2L2 # 3 смисъл CACCGGAGTAGTTGGCAGATCCAC, антисенс AAACGTGGATCTGCCAACTACTCC) бяха закалени и се лигира в BsmB1 намали LentiCRISPR / eCas9 1.1. Лентивирусно инфектираните RPE-1 клетки бяха избрани с пуромицин и поддържани като сборна популация. Нокаутът беше потвърден чрез имунофлуоресценция и Western blotting.
Клетъчна култура и трансфекции
Човешки пигментни епителни клетки на ретината, трансформирани с теломераза (RPE-1) (ATCC), бяха култивирани в Dulbecco's Modified Eagle Medium (DMEM), съдържаща 1 g/L глюкоза, допълнена с пеницилин, стрептомицин, 1X коктейл от неесенциални аминокиселини (Life Technologies), и 10% фетален говежди серум (Life Technologies). За siRNA-трансфекции, 30,000 35,000 х 10 0.3 клетки/mL бяха засети за една нощ. Клетките получават 1 nM siRNA, разредена в DMEM без серум и комбинирана с 2% интерферинов трансфекционен реагент (PolyPlus). За сенсибилизация на апоптоза клетките получават 3 nM Bcl-XL siRNA. Клетките бяха събрани XNUMX дни след трансфекцията.
Химикали, антитела и siRNA олиго
Антитела срещу ?-тубулин (Cell Signaling), ?-тубулин (Sigma), Drp1 (BD Biosciences), KEAP1 (Proteintech), Lamin B1 (Abcam), PARP (Cell Signaling), PMP70 (Abcam) и Tom20 (BD Biosciences ) се използват при разреждане 1:1000 за Western blotting и за имунофлуоресценция. Вътрешно анти-Nrf2 заешко антитяло се използва при 1:2000 за Western blotting [34], [59]. Сулфорафан (Sigma) и ставроспорин (Tocris) се използват съответно при 50 цМ и 1 цМ. siRNAs срещу Drp1 (Dharmacon), Nrf2 (Dharmacon), KEAP1 (Cell Signaling) и Bcl-XL (Cell Signaling) бяха използвани при 10 nM, освен ако не е отбелязано друго.
Имунофлуоресценция и in Vivo етикетиране
Клетките, засадени върху 18 mm стъклени покривни стъкла, бяха третирани с носител или лекарство, фиксирани в 3.7% формалдехид и след това пропускливи в 0.2% Triton X-100/PBS върху лед за 10 минути. Първичните антитела се инкубират в 3% говежди серумен албумин (BSA) в PBS за една нощ при 4 °C. След промиване с PBS, клетките се инкубират за 1 час в подходящи за видовете, Alexa488- или Alexa546-, конюгирани вторични антитела (разредени 1:1000) и 0.1 ug/mL DAPI (Sigma) в 3% BSA/PBS. Митохондриите се визуализират или чрез анти-Tom20 имунофлуоресценция, или чрез инкубиране на клетки в 200 nM MitoTracker Red CMXRos (Molecular Probes, Inc.) в DMEM без серум за 30 минути при 37 °C преди фиксиране.
Микроскопия и анализ на изображението
Имунофлуоресцентните проби се разглеждат на конфокален микроскоп LSM710 (Carl Zeiss). Микроснимките бяха заснети с помощта на обективи с маслено потапяне 63X или 100X и изображенията, коригирани и подобрени с помощта на Adobe Photoshop CS6. Анализът на съвместната локализация беше извършен с помощта на функцията за съвместна локализация на Carl Zeiss LSM710 с прагове, зададени ръчно, докато не се виждаше идентичността на пробите. Мащабните ленти навсякъде, освен ако не е посочено друго, са 10 m. Митохондриалната морфология беше оценена чрез заслепено оценяване. Ако митохондриите на клетката се поддържат като множествена, кръгла, дискриминираща точка, клетката се оценява като „деляне“. Ако отделните митохондрии са неразличими и цялата митохондриална мрежа изглежда непрекъсната, клетката се оценява като „сливане“. Всички други клетки, включително тези с групирани митохондрии, бяха оценени като „междинни“.
Субклетъчни фракционации
RPE-1 клетките се отглеждат до сливане. След промиване с PBS, клетките се подлагат на центрофугиране при 600 µg в продължение на 10 минути и се ресуспендират в 600 µL изолационен буфер (210 mM манитол, 70 mM захароза, 5 mM MOPS, 1 mM EDTA pH 7.4+1 mM PMSF). Суспензията се лизира 30 пъти в Dounce хомогенизатор. Фракция от хомогената се запазва като "цялоклетъчен лизат." Остатъкът се подлага на центрофугиране при 800°g в продължение на 10 минути до пелетни ядра. Супернатантите се подлагат на центрофугиране при 1500°g за 10 минути, за да се изчистят останалите ядра и нелизирани клетки. Този супернатант се подлага на центрофугиране при 15,000 15 ºg за XNUMX минути до пелетни митохондрии. Супернатантата се запазва като „цитозолна фракция“. Пелетата се промива внимателно с PBS и се ресуспендира в изолационен буфер. Концентрацията на протеин на всяка фракция се измерва чрез анализ на бицинхонинова киселина (BCA) и еквивалентните количества протеин се разделят чрез SDS-PAGE.
Western Blotting
Клетките се промиват в PBS и се разтварят в 2 пъти концентриран Laemmli солюбилизиращ буфер (100 mM Tris [pH 6.8], 2% SDS, 0.008% бромофенолов синьо, 2% 2-меркаптоетанол, 26.3% глицерол Y.0.001% PyO). Лизатите се варят в продължение на 5 минути преди зареждане върху полиакриламидни гелове на натриев додецил сулфат (SDS). Протеините се прехвърлят към нитроцелулозни мембрани и мембраните се блокират за 1 час в 5% мляко/TBST. Първичните антитела се разреждат в 5% мляко/TBST и се инкубират с блота за една нощ при 4 °С. Конюгирани с пероксидаза от хрян (HRP) вторични антитела се разреждат в 5% мляко/TBST. Блотовете бяха обработени с повишена хемилуминесценция и денситометричните количествени оценки бяха извършени с помощта на софтуер ImageJ.
Сулфорафанът е химикал от колекцията от изотиоцианат от органосерни вещества, получени от кръстоцветни зеленчуци, включително броколи, зеле, карфиол, къдраво зеле и кисели зеле, наред с други. Сулфорафанът се произвежда, когато ензимът мирозиназа трансформира глюкорафанина, глюкозинолат, в сулфорафан, известен също като сулфорафан-глюкозинолат. Кълновете от броколи и карфиолът имат най-висока концентрация на глюкорафанин или предшественик на сулфорафан. Изследванията показват, че сулфорафанът подобрява антиоксидантните способности на човешкото тяло за предотвратяване на различни здравословни проблеми. Д-р Алекс Хименес DC, CCST Insight
Сулфорафан и неговите ефекти върху рака, смъртността, стареенето, мозъка и поведението, сърдечните заболявания и др.
Изотиоцианатите са едни от най-важните растителни съединения, които можете да получите във вашата диета. В това видео правя най-изчерпателния случай за тях, който някога е бил направен. Кратко внимание? Прескочи до любимата си тема, като кликнете върху някоя от точките по-долу. Пълна времева линия по-долу.
Ключови секции:
00: 01: 14 - Рак и смъртност
00: 19: 04 - Стареене
00: 26: 30 - Мозък и поведение
00: 38: 06 - Краен рекап
00: 40: 27 - Доза
Пълна времева линия:
00: 00: 34 - Въвеждане на сулфорафан, основен фокус на видеото.
00: 01: 14 - Консумация на кръстоцветни зеленчуци и намаляване на смъртността от всякаква причина.
00: 02: 12 - риск от рак на простатата.
00: 02: 23 - риск от рак на пикочния мехур.
00: 02: 34 - Рак на белия дроб при риск от пушене.
00: 02: 48 - риск от рак на гърдата.
00: 03: 13 - Хипотетично: какво ще стане, ако вече имате рак? (Интервенция)
00: 03: 35 - Вероятен механизъм, който управлява асоциативните данни за рака и смъртността.
00: 04: 38 - Сулфорафан и рак.
00: 05: 32 - доказателства при животни, показващи силен ефект на екстракт от кълнове от броколи върху развитието на тумор на пикочния мехур при плъхове.
00: 06: 06 - Ефект на директно добавяне на сулфорафан при пациенти с рак на простатата.
00: 07: 09 - Биоакумулиране на изотиоцианатни метаболити в действителната гръдна тъкан.
00: 08: 32 - Инхибиране на стволови клетки от рак на гърдата.
00: 08: 53 - История: Brassicas са установени със здравни качества дори в древния Рим.
00: 09: 16 - Способността на Sulforaphane да подобри екскрецията на канцероген (бензен, акролеин).
00: 09: 51 - NRF2 като генетичен превключвател чрез елементи на антиоксидантен отговор.
00: 10: 10 - Как активирането на NRF2 подобрява екскрецията на канцероген чрез глутатион-S-конюгати.
00: 10: 34 - Брюкселското зеле увеличава глутатион-8-трансферазата и намалява увреждането на ДНК.
00: 11: 20 - Напитката от кълнове от броколи повишава екскрецията на бензен с 61%.
00: 15: 45 - Кръстосана консумация на зеленчуци и смъртност от сърдечни заболявания.
00: 16: 55 - Броколи зеленчуков прах подобрява кръвните липиди и цялостния риск от сърдечни заболявания при диабетици тип 2.
00: 19: 04 - начало на стареене секция.
00: 19: 21 - Диета, обогатена със сулфарафан, увеличава живота на бръмбарите от 15 до 30% (при определени условия).
00: 20: 34 - Значение на ниското възпаление за дълголетие.
00: 22: 05 - Кръстоцветните зеленчуци и кълнове от кълнове от броколи изглежда намаляват голямо разнообразие от възпалителни маркери при хората.
00: 23: 40 - Средно видеоклип: рак, стареене
00: 24: 14 - Изследванията на мишки показват, че сулфарафан може да подобри адаптивната имунна функция в напреднала възраст.
00: 25: 18 - Сулфарафан подобри растежа на косата при миши модел на оплешивяване. Изображение на 00: 26: 10.
00: 26: 30 - начало на секцията за мозъка и поведението.
00: 27: 18 - Ефект на екстракт от кълнове от броколи върху аутизма.
00: 27: 48 - Ефект на глюкорафанин върху шизофрения.
00: 28: 17 - Начало на депресивната дискусия (правдоподобен механизъм и проучвания).
00: 31: 21 - Изследване на мишки, използващи различни модели 10 на стресова индуцирана депресия, показва сулфорафан с подобен ефект като флуоксетин (prozac).
00: 32: 00 - Изследването показва, че директното поглъщане на глюкорафанин в мишки е подобно ефективно при предотвратяване на депресията от модела на социално потискане на стреса.
00: 33: 01 - начало на секцията за невродегенерация.
00: 33: 30 - Сулфорафан и болестта на Алцхаймер.
00: 33: 44 - Сулфорафан и болестта на Паркинсон.
00: 33: 51 - Сулфорафан и болестта на Хънтингтън.
00: 34: 13 - Сулфорафан увеличава протеините на топлинния шок.
00: 34: 43 - Начало на травматичното мозъчно увреждане.
00: 35: 01 - Сулфарафан се инжектира веднага след като TBI подобрява паметта (изследване на мишката).
00: 35: 55 - Сулфорафан и невронна пластичност.
00: 36: 32 - Сулфорафан подобрява ученето при модел на диабет тип II при мишки.
00: 37: 19 - Сулфорафан и душевна мускулна дистрофия.
00: 37: 44 - Инхибиране на миостатина в мускулните сателитни клетки (in vitro).
00: 38: 06 - Късно видеоклип: смъртност и рак, увреждане на ДНК, оксидативен стрес и възпаление, екскреция на бензол, сърдечно-съдови заболявания, диабет тип II, ефекти върху мозъка (депресия, аутизъм, шизофрения, невродегенерация).
00: 40: 27 - Мисли за определяне на доза от кълнове от броколи или сулфорафан.
00: 41: 01 - Анекдоти за поникване у дома.
00: 43: 14 - На температурата на готвене и активността на сулфорафан.
00: 43: 45 - Конверсия на сулфорафан от глюкорафанин в чревната бактерия.
00: 44: 24 - Добавките работят по-добре, когато се комбинират с активна мирозиназа от зеленчуци.
00: 44: 56 - техники за готвене и кръстоцветни зеленчуци.
Нагряването намалява активността на епитиоспецифичния протеин и увеличава образуването на сулфорафан в броколите
абстрактен
Сулфорафанът, изотиоцианат от броколи, е един от най-мощните антиканцерогени, получени от храни. Това съединение не присъства в непокътнатия зеленчук, по-скоро се образува от неговия глюкозинолатен предшественик, глюкорафанин, чрез действието на мирозиназа, тиоглюкозидазен ензим, когато тъканта на броколи се смачква или дъвче. Въпреки това, редица проучвания показват, че добивът на сулфорафан от глюкорафанин е нисък и че небиоактивният нитрилен аналог, сулфорафан нитрил, е основният продукт на хидролиза, когато растителната тъкан се раздробява при стайна температура. Последните данни показват, че при Arabidopsis образуването на нитрили от глюкозинолатите се контролира от топлочувствителен протеин, епитиоспецифичен протеин (ESP), некаталитичен кофактор на мирозиназа. Нашите цели бяха да проучим ефектите от нагряване на цветчета и кълнове на броколи върху образуването на сулфорафан и сулфорафан нитрил, да определим дали броколите съдържат ESP активност, след това да съпоставим зависими от топлината промени в ESP активността, съдържанието на сулфорафан и биоактивността, измерени чрез индукция на фаза II детоксикационен ензим хинон редуктаза (QR) в клетъчна култура. Загряването на пресни цветчета от броколи или кълнове от броколи до 60 °C преди хомогенизирането едновременно увеличава образуването на сулфорафан и намалява образуването на сулфорафан нитрил. Значителна загуба на ESP активност успоредно с намаляването на образуването на сулфорафан нитрил. Загряването до 70 °C и повече намалява образуването на двата продукта в цветчетата броколи, но не и в кълновете от броколи. Индукцията на QR в култивирани миши хепатомни Hepa lclc7 клетки паралелно увеличава образуването на сулфорафан.
Предварителното загряване на цветчетата и кълновете на броколи до 60 °C значително повишава катализираното от мирозиназа образуване на сулфорафан (SF) в екстракти от растителна тъкан след раздробяване. Това е свързано с намаляване на образуването на сулфорафан нитрил (SF нитрил) и активността на епитиоспецифичен протеин (ESP).
В заключение, сулфорафанът е фитохимикал, открит в броколите и други кръстоцветни зеленчуци. Неконтролирано количество оксиданти, причинено както от вътрешни, така и от външни фактори, може да причини оксидативен стрес в човешкото тяло, което в крайна сметка може да доведе до различни здравословни проблеми. Сулфорафанът може да активира производството на Nrf2, транскрипционен фактор, който помага за регулиране на защитните антиоксидантни механизми, които контролират реакцията на клетката към оксиданти. Обхватът на нашата информация е ограничен до хиропрактика и проблеми със здравето на гръбначния стълб. За да обсъдите темата, моля, не се колебайте да попитате д-р Хименес или да се свържете с нас на 915-850-0900 .
Допълнителна дискусия на тема: „Остра болка в гърба
Заболявания на опорно-двигателния апаратЕ една от най-разпространените причини за увреждане и пропуснати дни на работа в световен мащаб. Болките в гърба се дължат на втората най-честа причина за посещения в лекарски кабинет, превъзхождани само от инфекции на горните дихателни пътища. Приблизително 80% от населението ще изпитва болки в гърба поне веднъж през живота си. Гръбначният стълб е сложна структура, съставена от кости, стави, връзки и мускули, наред с други меки тъкани. Поради това наранявания и / или влошени състояния катохерния дискове, може евентуално да доведе до симптоми на болки в гърба. Спортни наранявания или наранявания при автомобилни аварии често са най-честата причина за болки в гърба, но понякога най-простите от движенията могат да имат болезнени резултати. За щастие алтернативните възможности за лечение, като хиропрактика, могат да помогнат за облекчаване на болката в гърба чрез използване на корекции на гръбначния стълб и ръчни манипулации, като в крайна сметка подобряват облекчаването на болката.
Инструментът Find A Practitioner на IFM е най-голямата мрежа за препращане във функционалната медицина, създадена, за да помогне на пациентите да намерят практикуващи функционална медицина навсякъде по света. Сертифицираните практикуващи от IFM са изброени на първо място в резултатите от търсенето, като се има предвид тяхното обширно образование по функционална медицина